编译:微科盟承乐,编辑:微科盟居居、江舜尧。
微科盟原创微文,欢迎转发转载,转载须注明来源【微生态】公众号。
导读
尽管大气纳米塑料(NPs)越来越普遍,但对其植物毒性、叶片吸收和在植物中易位的研究仍然有限。本文旨在通过研究番茄叶片暴露于NPs(带不同电荷)的生理效应以及NPs的叶片吸收和易位来填补这一知识空白。本研究发现,带正电荷的NPs会引起更显著的生理效应,包括生长抑制、抗氧化酶活性增加、基因表达和代谢物组成改变,甚至显著改变了叶际微生物群落的结构和组成。此外,带不同电荷的NPs表现出不同的叶片吸收和易位,带正电荷的NPs更多地渗透到叶片中,并在叶肉细胞内均匀分布。此外,叶片吸收的NPs能够易位到根部。这些发现为了解大气NPs与作物之间的相互作用提供了重要见解,并表明NPs在作物中的积累可能对农业生产和食品安全产生负面影响。

图文摘要
论文ID
原名: Toxicity Mechanisms of Nanoplastics on Crop Growth, Interference of Phyllosphere Microbes, and Evidence for Foliar Penetration and Translocation
译名: 纳米塑料对作物生长的毒性机制、叶际微生物的干扰以及叶渗透和易位的证据
期刊: Environmental Science & Technology
IF: 11.4
发表时间: 2023.11
通讯作者: 刘维涛,周启星,邢宝山
通讯作者单位: 南开大学环境科学与工程学院;美国马萨诸塞大学
DOI号: 10.1021/acs.est.3c03649
实验设计

结果与讨论
1 NPs暴露的物理-生化效应
本研究表明,叶片暴露于NPs降低了番茄的生长。此外,在PS-SO3H-H和PS-H处理下观察到萎黄病症状和疾病感染(图1a,f)。暴露于PS-SO3H的番茄植株鲜重没有显著下降。然而,当暴露于PS时,植株鲜重显著降低( p < 0.05),与对照组相比,分别降低41.7%(PS-L)和51.5%(PS-H)(图1b)。不同水平的NPs暴露对株高的影响有显著差异( p < 0.05)。在高剂量下,PS对株高的影响与PS-SO3H也有很大的差异( p < 0.05)(图1b)。除PS-SO3H-L外,NPs显著降低了叶片的叶绿素含量( p < 0.05)(图1c)。有趣的是,低剂量NPs(PSSO3H-L和PS-L)暴露下的净光合速率(Pn)、蒸腾速率(Tr)和气孔导度(Gs)增加,而高剂量NPs(PSSO3H-H和PS-H)暴露下则降低(图1d)。低剂量NPs对光合作用的刺激和高剂量NPs的抑制与纳米材料「毒物兴奋效应」现象中的剂量−反关系相一致。研究发现,低浓度的纳米颗粒可以通过增加叶绿体膜的渗透性来增强叶绿体希尔(Hill)反应活性并加速O2释放,从而使Ca2+和Cl−更容易进入放氧复合体的中心,从而促进叶绿体的光合磷酸化活性。
叶片暴露于NPs后的光合抑制被认为是影响植物生长的主要机制。NPs暴露会导致O2-和MDA含量显著增加。虽然SOD、POD、CAT等抗氧化酶的活性增加,但其清除作用并没有达到维持稳态所需的基线活性氧(ROS)水平。过量的ROS可能会破坏叶绿体结构,抑制光合作用。在暴露于NPs的叶片细胞中,叶绿体结构增大,嗜锇颗粒增加,证实了上述观点(图1e)。高剂量和低剂量NPs的影响之间存在显著差异,表明番茄抗氧化应激系统对NPs的叶片暴露水平非常敏感。当将拟南芥的根和玉米叶片暴露于NPs时也观察到了这种现象。对藻类的研究表明,附着在藻类细胞表面的NPs可以阻挡外部光,从而影响光合作用。这种物理效应也可能适用于高等植物。以往的研究发现,CuO和ZnO纳米颗粒可以吸附在植物表面,从而产生遮蔽作用,抑制植物的光合作用。此外,随后的研究发现,带正电荷的NPs(F-PS-Np)比带负电荷的NPs(F-PS-SO3H)在叶片表面吸附得更多。在表面涂层对植物纳米颗粒吸收影响的研究中也发现了类似的现象。这种差异主要归因于NPs与叶片表面的静电相互作用不同。带负电荷的NPs与带负电荷的细胞壁之间的静电排斥可导致积累减少,而带正电荷的NPs由于静电吸引而大量积累。

图1.NPs对番茄生长的影响。NPs对植物形态的影响(a);植株鲜重和株高(b)。NPs对光合作用的影响:叶绿素含量(c);光合作用参数(d);叶片形态(f);CK、PS-SO3H-H和PS-H处理组叶片的超微结构图(e)。(a)和(f)的比例尺分别是8 cm和6 mm。(b)和(c)中的数据以平均值±SD表示(n=3);不同的小写字母表示不同处理之间差异显著( p <0.05)。CK:对照组;PS-SO3H-L/PS-L:番茄植株叶片暴露于0.45 μg d−1带负电荷/正电荷的NPs下21天;PS-SO3H-H/PS-H:番茄植株叶片暴露于4.5 μg d−1带负电荷/正电荷的NPs下21天。
2 番茄多组学分析
为了研究上述效应的潜在机制,本研究对PS-SO3H-H和PS-H处理组的番茄叶片进行了转录组学和代谢组学分析(从此处开始,统一描述为PS-SO3H和PS)。PS对基因调控的作用强于PS-SO3H(图2a)。为了进一步研究转录组变化,本研究用GO和KEGG对DEGs进行了功能注释。PS-SO3H和PS均上调了MAPK信号通路,这是由NPs暴露后的细菌性斑点病感染引起的(图1f)。MAPK级联反应在感知病原体信号分子时被激活,以增强植物对疾病的抗性。此外,NPs还调节植物-病原体的相互作用通路。这一观察结果与Wang等人的类似结果一致,他们报道了PS-MPs诱导生菜叶片中与抗病通路相关的基因下调。
除上述几方面外,NPs还可以通过干扰光合色素和光合位点的产生来影响植物的光合作用。当暴露于PS-SO3H时,类胡萝卜素的生物合成过程明显下调,这影响了叶片的光吸收和抗氧化能力。GO分析也显示了类似的结果。PS-SO3H显著影响蛋白激酶活性和蛋白自磷酸化,这在植物抗逆性中起着关键作用,且后者是植物色素的重要来源。对于PS,包括叶绿体类囊体膜和质体类囊体膜在内的GO terms受到显著影响,这是光合作用的光反应位点。核酮糖-1,5-二磷酸羧化酶是碳固定的关键酶,其催化核酮糖-1,5-二磷酸生成3-磷酸甘油酸。在本研究中,PS暴露下调了编码二磷酸核酮糖羧化酶大亚基的 rbcL ,干扰了无机碳向有机化合物的转化,从而抑制了番茄植株的光合效率和生物量。
在代谢组学方面,通过主成分分析(PCA)和偏最小二乘判别分析(PLS-DA)发现,NPs暴露显著改变了叶片的代谢产物。差异代谢物(DM)主要包括碳水化合物、氨基酸和次生代谢物(图2b)。碳水化合物是植物生长发育所必需的能量来源。松二糖、乳糖、果糖和山梨糖含量的降低可能是由于NPs诱导的氧化应激,这会增加与能量代谢相关的糖降解。氨基酸是各种次生代谢产物的前体,在植物生理过程中起着至关重要的作用。丝氨酸和L-谷氨酸参与叶绿素合成,其含量降低会导致叶绿素含量降低(图1c)。缬氨酸和脯氨酸可作为抗氧化剂和信号分子参与植物对非生物胁迫的反应。PS暴露显著降低了缬氨酸和脯氨酸的水平,表明PS诱导了更多的氧化应激。
本研究对转录组学和代谢组学进行了相关网络分析,进一步探索番茄的调控网络。许多DEGs和DMs的相关性很强( p < 0.05,R > 0.9)(图2c)。这些DEGs主要富集在氨酰-tRNA生物合成、丁酸甲酯代谢、丙氨酸、天冬氨酸和谷氨酸代谢以及谷胱甘肽代谢通路的代谢物中(图2d)。谷胱甘肽代谢是植物在面临胁迫时维持关键生物过程的机制。此外,氨酰-tRNA由合成酶形成,是蛋白质合成的组成部分。NPs对氨酰-tRNA生物合成的破坏表明,需要关注作物通过调节翻译过程来应对NP胁迫的作用。
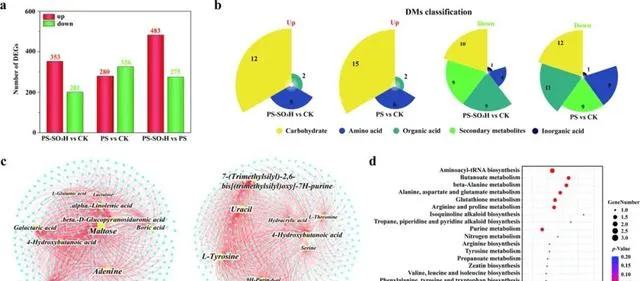
图2.番茄叶片施用NPs后的转录组学和代谢组学分析。番茄叶片的DEGs数量(a)和DMs的分类统计(b)。转录组和代谢组变化的关联分析。不同处理下的前10个DMs和DEGs之间的关系网络(|log2FC|≥2, p ≤0.05)(c);使用气泡图显示共现网络中代谢物的主要代谢通路富集结果(d)。在共现网络中,黄色和青色节点分别代表DMs和DEGs。节点的大小与连接到节点的边数有关。CK:对照组;PS-SO3H-H/PS-H:番茄植株叶片暴露于4.5 μg d−1带负电荷/正电荷的NPs下21天。
3 叶际微生物群落的变化
叶际是各种微生物的重要栖息地。叶际为微生物提供营养;微生物有助于植物的生长和发育。当面对外部环境胁迫时,叶际微生物种群可能会发生变化,从而影响它们与宿主的相互作用。
β多样性分析显示,各处理间的聚类位置显著分离,表明NPs对番茄叶际微生物群落结构有显著影响(图3a)。α多样性分析显示,与对照组相比,NPs暴露提高了Chao、Ace和Shannon指数。特别是,PS暴露更显著地改变了叶际微生物群落的多样性,并诱导了特定微生物种群的富集(图3b)。此外,MNPs也可以改变门水平细菌组成。这种变化通常是由能够生物降解MNPs的细菌丰度增加而引起的。在本研究中,NP处理组塑料降解菌(如变形菌门和蓝细菌门)的相对丰度增加(图3c)。在科水平上,芽孢杆菌科在番茄叶际中占优势,其在对照组中的相对丰度为76.43%,在PS-SO3H和PS暴露后分别降至56.85%和37.06%。芽孢杆菌是公认的有益菌,能够促进植物生长,抵御病原菌感染。NPs诱导的芽孢杆菌科相对丰度降低可能是番茄抗病性受到干扰的原因。
微生物通常会发展出复杂的网络来应对外部干扰,而核心微生物可以通过微生物之间的相互作用强烈地影响整个微生物群落的结构。丛毛单胞菌科(Comamonadaceae)是一个重要的微生物分类群,已被证明可以有效地控制叶际微生物丰度,其在NPs暴露后显著增加。本研究分析了番茄叶际微生物的共现网络。NPs处理干扰了微生物群落的网络复杂性(图3d)。其中,与对照组相比,NPs暴露后叶际微生物网络的边数、平均度、聚类系数和网络直径均显著增加。有研究表明,叶际微生物通过形成生物膜增加了MNPs在叶片表面的聚集。相应地,NPs在植物表面的沉积为微生物提供了新的生态位,从而为叶际微生物相互作用提供了机会。这有助于解释本研究中NPs增强了细菌之间的关系,并加强了微生物群落的共现模式。
植物叶际微生物相互作用在维持植物健康和抑制病原菌过度生长方面发挥着重要作用。它们可能受到多种生物因素(如叶片代谢物和植食性昆虫)和非生物因素(如气候因子)的影响。本研究还发现,大量的DMs与番茄叶际细菌门之间存在显著的相关性。含碳和含氮营养物质的可用性是限制附生微生物定植的主要因素。暴露于NPs后,番茄叶片代谢物中碳水化合物和氨基酸组分显著增加。以往的研究表明,叶片养分与叶际微生物群落的稳定性呈正相关。这与本研究结果相似,表明NPs可以通过改变植物代谢物来促进新的优势微生物群落的形成,并形成叶际微生物群落。
利用PICRUSt预测叶际微生物功能的变化。KEGG一级通路中富集三种通路(代谢、细胞过程和环境信息处理)。在二级通路中,有5条通路发生了显著改变,其中4条是代谢通路。因此,叶际微生物的代谢也受到了干扰。此外,PS暴露显著增加了细胞生长与死亡通路的丰度,从59.93%增加到69.58%。叶际微生物生理过程的提升可能是微生物网络高度复杂的原因。

图3.叶际微生物群落对NPs的响应。NMDS分析(a),对照组和NPs处理之间细菌α多样性(Chao、Ace和Shannon指数)比较(b)。细菌门和细菌科的相对丰度(c)。NPs对细菌群落共现模式的影响(d)。节点的大小根据节点的度进行缩放,并根据模块对节点进行着色。(b)中的数据以平均值±SD表示(n=3)。* p <0.05,** p <0.01。CK:对照组;PS-SO3H/PS:番茄植株叶片暴露于4.5 μg d−1带负电荷/正电荷的NPs下21天。
4 电荷控制NPs内化进入叶片
为了证明番茄叶片对NPs的吸附和吸收,本研究使用CLSM研究了NPs的转移效率和分布。根据荧光NPs稳定性测试结果,F-PS-SO3H和F-PS-Np NPs在模拟植物溶液和原生质体细胞液中24 h内均未检测到荧光释放。冲洗后,在4 h时,叶片表面的F-PS-Np荧光强度强于F-PS-SO3H,表明叶片表面对带正电荷的NPs的吸附能力更强。由于植物细胞壁带负电荷,这一发现是符合预期的。此外,本研究发现PS-SO3H与叶片的接触角高达105.80 ± 0.47,导致液滴呈球形,容易丢失。PS-Np与叶片的接触角为93.96 ± 0.20,液滴呈扁球形,更容易吸附在叶片上,并可快速扩散。这导致F-PS-Np在叶片表面呈均匀分布。同时,F-PS-SO3H大量聚集,进一步阻碍了NPs的内化。
本研究通过CLSM图像和3D视频显示了不同时间NPs在叶肉中的分布和传递情况。在CLSM图像中,观察到了更多的F-PS-Np,表明带正电荷的NPs比带负电荷的NPs更能有效地进入叶肉。随着时间的推移,两种NPs的转移效率都有所提高(图4)。F-PS-Np和F-PS-SO3H的分布也有显著差异。F-PS-Np均匀分布并被转移到叶肉细胞中。相比之下,F-PS-SO3H只有在暴露时间超过24 h后才会少量进入细胞内部(图4a)。
气孔和表皮被认为是叶片吸收NPs的两个主要途径。本研究发现,叶片可以通过这两种途径吸收带不同电荷的NPs。NPs可以通过质外体(胞外)和共质体(胞内)途径在叶片内易位。本研究发现,带正电荷的NPs在叶绿体中的转运途径是共质体,而带负电荷的NPs则通过质外体途径转运(图5)。后者更容易理解,因为双子叶植物的叶子同时包含薄膜状和海绵状的叶细胞。这些细胞排列稀疏,形成了较大的胞外空间,但NPs从细胞外空间进入叶肉细胞时会遇到细胞壁和细胞膜等屏障。
植物细胞壁被认为是阻挡纳米材料的主要屏障,其标准平均孔径小于10 nm,远远小于本研究中的NPs。虽然纳米颗粒的吸收和易位受到尺寸排斥的限制,但与本研究结果相似,据报道高达36−50 nm的工程纳米颗粒(ENPs)可以被内化和易位。尺寸排斥是动态的,并直接受到钙、蛋白质、病毒或ENPs的影响。常见的ENPs(如SiO2NPs和TiO2 NPs)可以诱导细胞壁形成新的或更大的孔隙,从而改变细胞壁结构,促进较大ENPs的内化。与这些ENPs相比,本研究中使用的NPs更柔软。本研究观察到被植物内化的NPs发生了变形(图7c),这有利于NPs的吸收和易位。Li等人也报道了类似的发现。
由于发现NPs已经进入植物细胞,本文研究了原生质体(无细胞壁)对NPs的摄取。本研究在健康的原生质体中发现了与完整叶片相同的积累模式,其中F-PS-SO3H聚集在原生质体周围,F-PS-Np在原生质体中聚集。与本研究结果相似,有报道称,带正电荷的颗粒比带负电荷的颗粒更容易被细胞吸附并通过内吞作用穿过细胞膜。然而,也有报道称,软塑料颗粒的表面正电荷抑制了它们穿透植物细胞膜的能力。由于软塑料的特异性,植物细胞可能通过不同于硬塑料的途径来吸收它们。例如ZnO和Au等硬颗粒可以进入原生质体,而与电荷无关。与带负电荷的粒子相比,带正电荷的粒子被快速内化,这与本研究结果相似。易位的差异可能与粒子的zeta电位有关。据报道,随着纳米颗粒zeta电位的增加,脂质双分子层诱导的电位下降也随之增加,这将导致膜张力的增加,从而导致纳米颗粒更强的易位。在本研究中,F-PS-Np的zeta电位绝对值高于F-PS-SO3H,表明这一因素可能会影响NPs的易位效率。
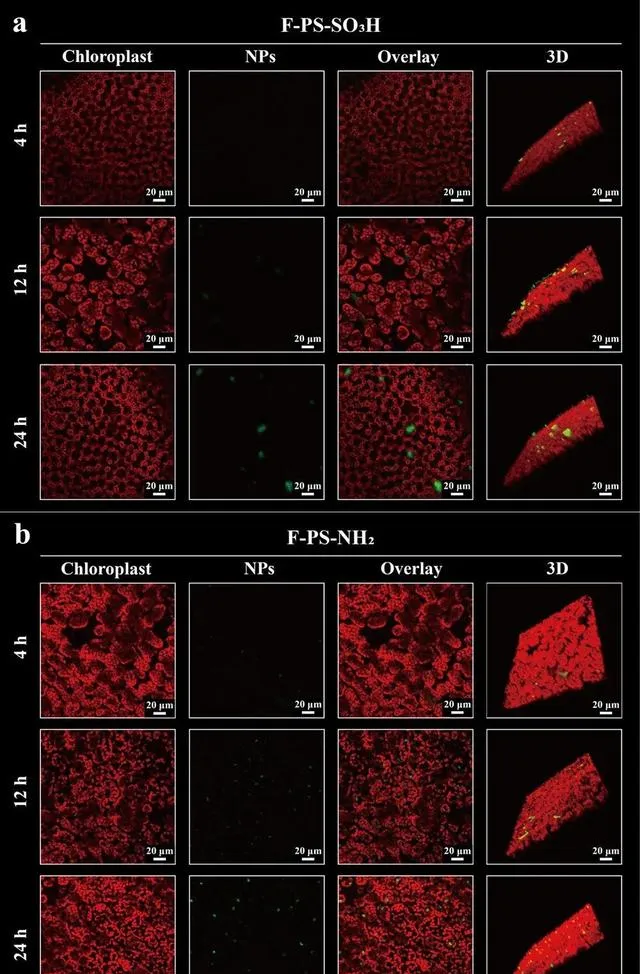
图4.叶片中NPs的易位和分布。4 h、12 h和24 h时叶肉中F-PS-SO3H (a)和F-PS-Np (b) NPs荧光强度的CLSM图像。番茄植株叶片暴露于F-PS-SO3H或F-PS-Np NPs(约2μg NPs/叶)。绿色是NP粒子,红色是叶绿体。
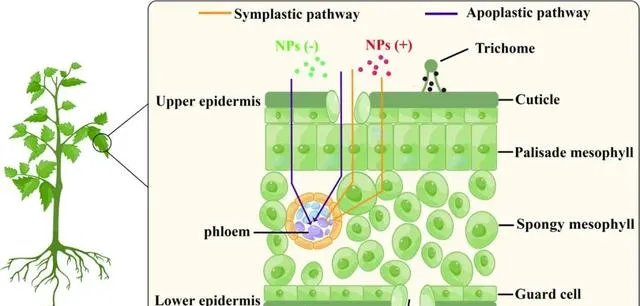
图5.番茄叶片中带不同电荷的NPs的易位途径及其分布。NPs通过气孔和/或表皮途径被吸收到叶片中,然后通过质外体途径(紫色线)在细胞外空间和细胞壁之间转移,或通过共质体途径(橙色线)进入叶肉细胞。表面电荷的差异影响NPs的易位途径。NP(−):带负电荷的NPs;NP(+):带正电荷的NPs。
5 NPs的叶片-根部易位及分布
为了进一步证实NPs的吸收和易位,本研究使用直径为70 nm的聚苯乙烯纳米塑料掺杂铕螯合物Eu-β-diketonate (PS-Eu),以定量番茄不同组织中的NPs。对于维管组织,韧皮部负责将光合产物从叶片长距离运输到其他组织。本研究发现,被叶片吸收的PS-Eu可以发生易位,但其易位能力有限。在第7天,内化到叶片中的PS-Eu只有3.51 ± 0.092%转移到根部,且随着施用剂量和时间的增加没有显著变化(图6b)。通过扫描电子显微镜(SEM)也观察到了同样的现象(图7)。很多PS-Eu颗粒在番茄叶片中聚集成簇(图6c,7a-f)。在根部只观察到少量的PS-Eu,其呈现为单独分散的颗粒或是聚集成簇(图6c,7g-l)。
造成这种现象的原因有很多,其中NPs的大小和聚集状态被认为是影响吸收和易位的主要因素。叶片吸收的大小限制可能取决于植物类型和NP特性等因素,因此确切的限值仍不清楚。一项研究表明,100 nm的PSNPs也可以被叶片吸收并转运到植物根部,其粒径比本研究选择的PS-Eu NPs大得多。NPs的聚集是抑制还是减少叶片吸收尚不清楚,但有报道称聚集的NPs被保留在叶片表面,这在本研究中也有发现。被叶片吸收的NPs在到达维管组织之前必须穿过几个屏障,包括表皮、质外体或共质体,这些都将进一步阻止NPs在叶片内的运输。施用于叶片的NPs到达韧皮部后,可通过由加宽的胞间连丝组成的筛管转运到植物的其他部位。NPs在韧皮部的易位过程中会遇到光合产物、螯合分子和不同的pH梯度。在这一过程中,NPs可能聚集、溶解或形成蛋白冠。因此,除了大小限制外,NPs的韧皮部转运非常复杂,其具体机制尚不清楚,仍有待进一步探索。
本研究在405 nm激发下,测定了PS-Eu处理7 d的番茄幼苗。在番茄根、茎和叶中观察到蓝色荧光(Eu的自身荧光)。本研究还发现,蓝色荧光主要集中在茎的韧皮部,这是NPs在韧皮部运输的直接证据。毛状体被认为是工程纳米颗粒进入叶片的可能途径。有趣的是,本研究也在叶片的毛状体中发现了PS-Eu。然而,这些NPs是被毛状体直接内化,还是被表皮内化后转移到毛状体内,仍有待进一步探索。此外,本研究在番茄茎的毛状体内部和外部都发现了PS-Eu,即使它们没有直接暴露于PS-Eu。PS-Eu很可能通过毛状体特定部位的组织间隙被挤压出来,这可能是植物通过自身排泄来解毒的主要机制。
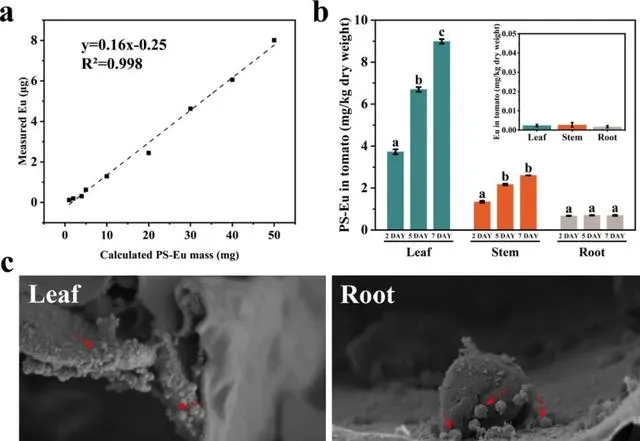
图6. PS-Eu在番茄中的分布。PS-Eu计算质量与Eu之间的相关性(a)。2、5和7天时PS-Eu NPs在番茄叶片、茎和根中的生物积累(b)。右上方的图显示了未暴露PS-Eu植株不同组织中Eu的含量。(a)和(b)中的数据以平均值±SD表示(n=3)。不同字母表示处理天数之间存在显著差异( p <0.05)。PS-Eu在番茄叶片和根中的分布(c)。番茄植株叶片连续7天暴露于2 μg PS-Eu NPs/leaf。实心箭头表示PS-Eu。

图7. PS-Eu在番茄叶片(a-f)和根部(g-l)分布的SEM图像。番茄植株叶片连续7天暴露于2 μg PS-Eu NPs/leaf。每个虚线箭头表示红框内区域的放大视图。实心箭头表示PS-Eu粒子(c,f,i和l)。
结论
综上所述,NPs可以进入番茄叶片,但带不同电荷的NPs在叶片吸收和分布方面存在显著差异。正电荷有利于NPs进入叶片并在叶片内均匀分布。NPs的细胞摄取也观察到类似的结果。NPs在番茄组织中的积累和分布对植物生长有不同的影响,带正电荷的NPs比带负电荷的NPs对植株生长的抑制更严重。此外,积累在叶片中的NPs可以通过韧皮部运输到植物根部。未来需要注意的是,在叶片中积累的NPs是否会转移到可食用的果实部分,从而对人类健康造成潜在危害。NPs在韧皮部的转运机制尚不清楚。有必要利用蛋白质组学等方法来探索可能参与NPs韧皮部运输的重要蛋白质。总体而言,本研究有助于更好地理解NPs与作物叶片的相互作用,这有助于了解NPs的归趋和影响。
然而,受检测技术和取样困难的限制,只有少量研究调查了大气中NPs的含量、组成和形态。为了获得更真实的环境NPs水平的影响,随后应尝试收集灰尘和蔬菜样本,并确定NPs的数量和类型,以进行进一步的研究。此外,NPs还会经历风化、分解等过程,从而导致这些NPs的理化性质发生变化。因此,有必要在更实际的环境下探讨风化NPs对作物生长和营养质量的影响。











