編譯:微科盟君影,編輯:微科盟居居、江舜堯。
微科盟原創微文,歡迎轉發轉載,轉載須註明來源【微生態】公眾號。
導讀
生產者的多樣性-功能關系以及這些關系如何受到消費者的影響是眾所周知的。然而,土壤中的微生物分解者並不為人所知。大型哺乳食草動物等消費者是否以及如何影響土壤微生物分解者多樣性-功能關系也仍然未知。本研究進行了一項為期14年的食草動物排除實驗,以研究微生物功能是否隨微生物多樣性(物種豐富度和組成)而變化,以及食草動物是否改變了多樣性-功能關系。在2019年的生長季節,對放牧和圍欄地塊的土壤進行了三次分析。結果表明,功能多樣性與微生物群落組成呈正相關,但與物種豐富度不相關;它也不受大型哺乳食草動物排除的影響,這表明它具有恢復力和抗性。群落組成和功能多樣性之間的這種正相關關系挑戰了超多樣化土壤微生物群落中普遍存在的功能冗余概念,因為某些物種組合可能優於其他物種,並決定分解過程和服務。結構方程式模型表明,這種關系的強度受到土壤水分有效性的影響。微生物功能隨時間變量(如季節性)的變化強於隨空間變量(如土壤質地和pH值等土壤因子)的變化。雖然結果可能會受到研究功能的種類和數量的限制,但一旦有16個或更多的功能被量化,這種關系是可推廣和穩健的。旱地的分解可能特別容易受到微生物物種身份(而非物種數量)如何在持續和預測的氣候變遷下對不斷上升的降雨變化作出反應的影響。
論文ID
原名: Community composition, and not species richness of microbes, influences decomposer functional diversity in soil
譯名: 微生物群落組成(而非物種豐富度)影響土壤中分解者的功能多樣性
期刊: Soil Biology and Biochemistry
IF: 9.7
發表時間: 2023.10
通訊作者: Sumanta Bagchi
通訊作者單位: 印度科學學院
DOI號: 10.1016/j.soilbio.2023.109225
實驗設計
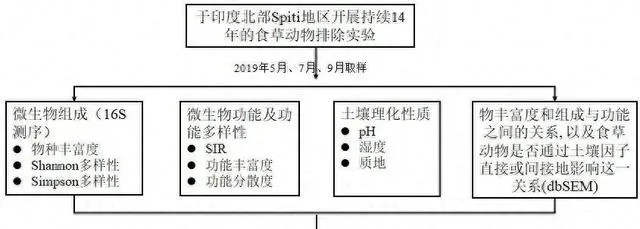
結果
1 微生物群落和功能
將7650894條reads聚類為47個已知細菌門的1937個獨特OTUs。其中,鑒定到屬水平的OTUs僅有924個。放線菌門、酸桿菌門、浮黴菌門、變形菌門和疣微菌門最為常見(圖1,圖S2)。每個樣本OTUs物種豐富度在494至866之間(平均713±80 SD,圖2)。Shannon多樣性在3.72至5.06之間(4.49±0.21,圖2),而Simpson多樣性在0.908至0.985之間(0.969±0.011,圖2)。同樣,功能豐富度(即代謝受質的數量)範圍在6到30之間(平均21±7 SD,圖1,圖S2)。
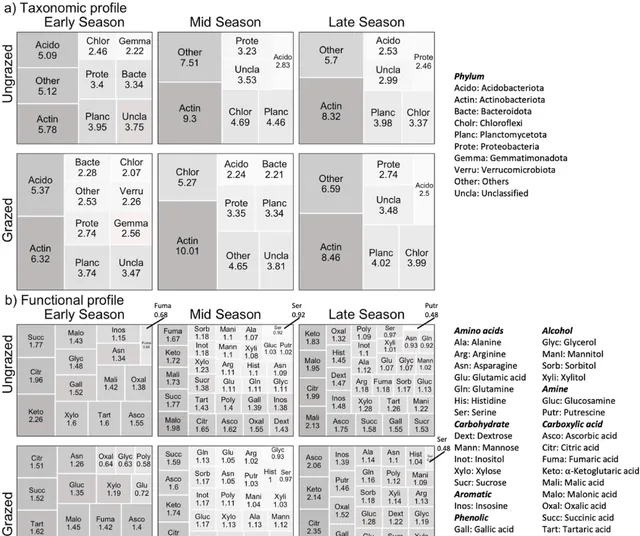
圖1.在有/無大型哺乳食草動物的情況下,所有地塊和整個生長季節(早、中、晚)的微生物分類譜(a, 門水平)和微生物功能譜(b,n = 30個受質)。每個單元格的面積分別與每個門的總豐度和每種受質的分解代謝活性成正比。數碼表示相對豐度(分類)和相對微生物活性(功能)的百分比。
2 季節性和食草動物對微生物種類和功能的影響
物種(OTUs)豐富度的變化既不能透過季節來解釋(圖2a;F2,45=0.02, P =0.98),也不能透過放牧來解釋(圖2a;F1,45=1.61, P =0.21)。Shannon多樣性隨放牧而變化(圖2b; F1,45 = 7.56, P = 0.01),但不隨季節而變化(圖2b; F2,45 = 0.03, P = 0.97)。Simpson多樣性不隨季節而變化(圖2c; F2,45 = 0.60, P = 0.56),也不隨放牧而變化(圖2c; F1,45 = 2.30, P = 0.13)。微生物功能豐富度隨季節而變化(圖2b;F2,45=38.96, P <0.001),但不隨放牧而變化(圖2b;F1,45=0.01, P =0.098)。同樣,微生物功能分散度隨季節變化(圖2c;F2,45=48.89, P <0.001),但不隨放牧而變化(圖2c;F1,45=0.01, P =0.092)。PCoA分析顯示,微生物物種組成隨季節而變化(圖3;F2,54=4.70, P =0.001),但與放牧無關(圖3;F1,54=1.33, P =0.21)。功能組成也隨季節變化(圖3,F2,54=4.83, P =0.001),但與放牧無關(圖3:F1,54=0.87, P =0.55)。
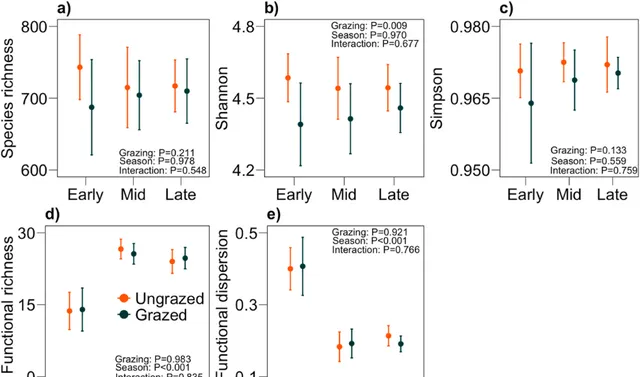
圖2.在整個生長季節有/無大型哺乳食草動物的情況下,土壤微生物的物種豐富度(a)、Shannon多樣性(b)、Simpson多樣性(c),功能豐富度(d)和功能分散度(e)。物種豐富度在放牧地和未放牧地之間以及生長季節之間無顯著差異。功能豐富度和功能分散度在放牧地和未放牧地之間沒有差異,但在生長季節之間存在差異。
圖3.主座標分析(PCoA)顯示在有/無大型哺乳食草動物的情況下以及生長季節不同時間點微生物群落和分解者功能的排序。總體而言,土壤微生物分解者的分類和功能組成在放牧地和未放牧地之間沒有變化,但在生長季節之間存在差異。
3 物種組成和功能之間的關系
微生物功能Kriged圖表明,群落組成差異與功能差異有關(圖4),並使用dbSEM進一步評估(圖5)。總體而言,dbSEM的功能豐富度和功能分散度(圖5)表明數據與假設路徑吻合良好(SRMR=0.005,RMSEA<0.001,P-value(RMSEA)=0.982,CFI>0.999,X2=1.34,P-value(X2)=0.512)。路徑系數表明,物種組成與功能豐富度呈正相關(物種組成→功能豐富度;路徑系數=0.08, P =0.06;圖5),但不含功能組成(物種組成→功能組成; P > 0.10;圖5)。物種豐富度對功能豐富度和功能組成均無直接或間接影響( P >0.10;圖5),土壤質地對物種組成有正向影響(質地→物種組成;路徑系數=0.09, P =0.001;圖5)。季節性與物種組成(季節→物種組成;路徑系數=0.14, P <0.001;圖5)、功能組成(季節→功能組成;路徑系數=0.25, P <0.001;圖5)、土壤濕度(季節→濕度;路徑系數=0.29, P <0.001;圖5)呈正相關,因此,隨著季節從季初到季中到季末的發展,物種組成、功能組成和土壤濕度的差異越來越大。但季節性與物種豐富度呈負相關(季節→物種豐富度;路徑系數=-0.04, P =0.08;圖5),因此隨著季節從季初到季中到季末的發展,樣品之間的物種豐富度趨於相似。由此可見,季節性對多樣性-功能關系既有直接影響(季節→物種組成→功能),也透過濕度(季節→濕度→物種組成→功能)間接影響。濕度的變化影響物種組成(濕度→物種組成;路徑系數=0.12, P =0.02;圖5)和物種豐富度(濕度→物種豐富度;路徑系數=0.08, P =0.09;圖5)。土壤pH值變化與物種豐富度呈負相關(pH→物種豐富度;路徑系數=-0.07, P =0.09;圖5)。放牧不直接影響這種關系(放牧→物種組成→功能),也不透過土壤因子間接影響這種關系(放牧→土壤→物種組成→功能)。
透過對功能數據進行叠代重采樣的思維實驗,發現16種或更多不同組合的受質可以提供功能多樣性的穩健解釋(圖6)。只要有16種或更多功能的定量資訊,就可以推斷出物種組成與功能之間的關系(圖5)。
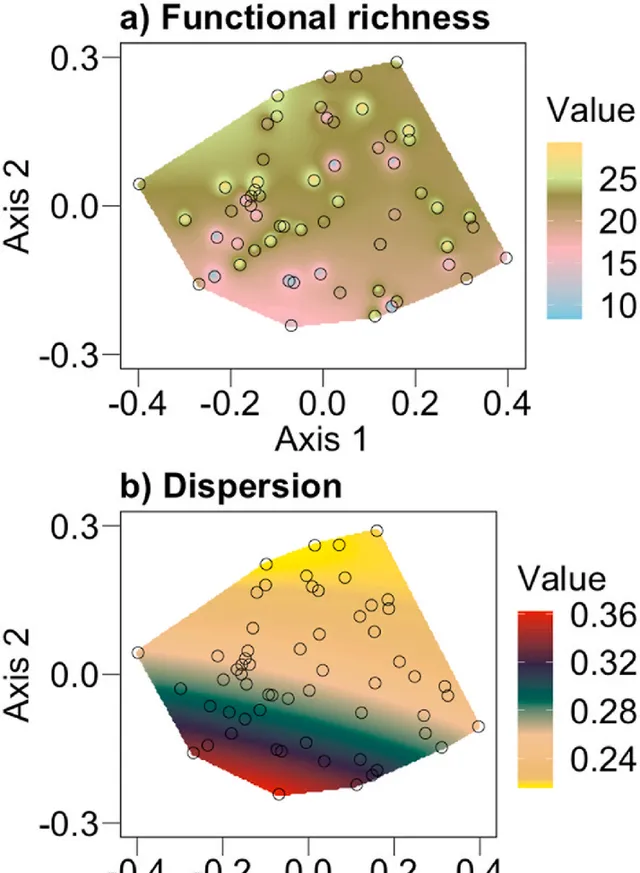
圖4. Kriged圖總結了微生物功能豐富度(a)和功能分散度(b)與微生物物種組成的關系,表明微生物分解者的功能可以隨物種組成而變化。彩色區域強調了在生長季節不同時間點測量的研究地點的微生物物種組成範圍內的微生物功能插值。物種組成用主座標分析(PCoA)的第一軸和第二軸表示。圓圈表示物種組成的采樣數據點。
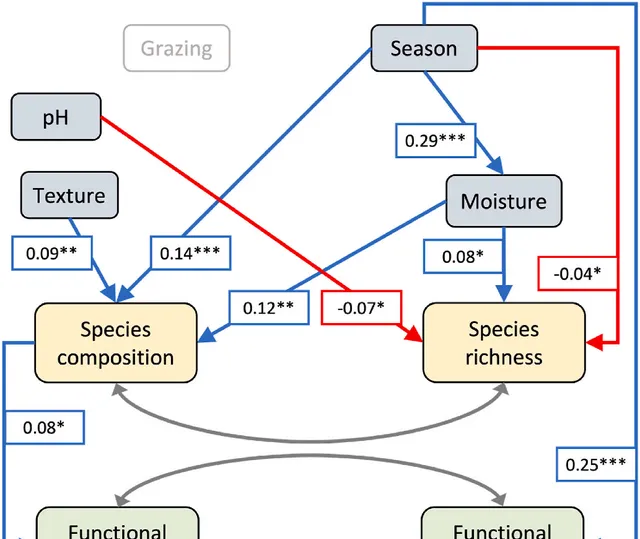
圖5.基於距離的結構方程式模型(dbSEM)評估放牧、季節、土壤非生物特性對物種多樣性與微生物功能關系的影響。數值表示標準化路徑系數,星號表示統計學顯著性(***P≤0.01, **P≤0.05, *P≤0.10)。放牧(放牧/未放牧)的模型路徑不顯著,因此未顯示。模型診斷為SRMR=0.004,RMSEA<0.001,P-value(RMSEA)=0.982,CFI>0.999,X2=1.34,df=2,P-value(X2)=0.512。總體而言,微生物功能與物種組成呈正相關,而物種豐富度對功能沒有影響。季節性透過土壤濕度的變化對這一關系有直接和間接影響。
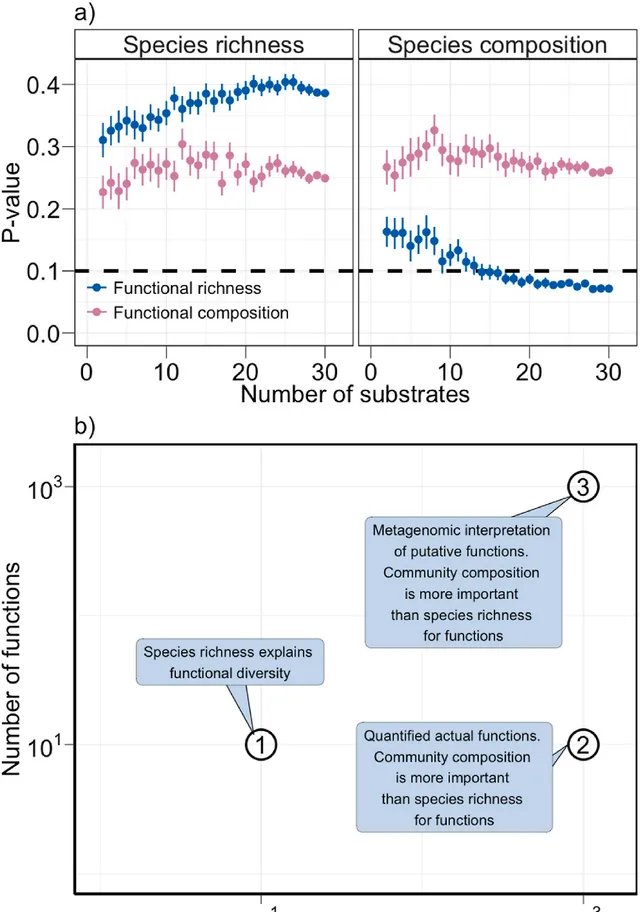
圖6.思維實驗總結,評估測量功能的數量如何影響多樣性-功能關系的解釋(a)。透過對功能數據進行叠代重采樣,生成了2800個子集,其中包含2-29個功能的100種不同的可能組合。對於每個子集,重復進行dbSEM分析(圖5),並檢查了連線物種多樣性(物種豐富度和物種組成)與功能(功能豐富度和功能組成)的路徑的顯著性(α=0.1)。報告了上述路徑的P值(物種多樣性→功能)。關於適應不同數量物種和功能的微生物多樣性-功能關系的三類研究的概念總結(b)。對101個物種和101種功能的實驗組裝群落的研究,通常會遇到物種豐富度和功能之間的正相關關系(即①)。對103個物種的超多樣化自然群落的研究,通常會遇到部份冗余(即②)。對大量假定功能進行宏基因組解釋的研究也會遇到部份冗余(即③)。思維實驗和概念總結進一步強調了微生物物種身份(而非物種數量)對理解土壤分解過程的重要性。
討論
雖然生產者的多樣性-功能關系以及它們在較高營養水平上受到消費者的調節是眾所周知的,但對於微生物分解者和分解功能來說,它們仍然是不確定的。本文研究了微生物分類多樣性是否與微生物分解功能相關,並行現了三個關鍵結果:(i)土壤微生物群落和功能隨時間的變化比隨空間的變化更大(圖1-3);(ii)微生物物種組成影響功能豐富度,但不影響功能組成(圖4-5,圖S4)。(iii)與土壤背景變量和季節性不同,大型哺乳食草動物不會影響微生物的多樣性-功能關系(圖5)。此外,覆蓋一系列分解者代謝過程的一小部份功能(16種或更多與碳代謝相關的受質)提供了足夠的樣本量來理解土壤微生物的多樣性-功能關系(圖6)。
本研究發現,微生物群落結構和功能的時間變化比空間異質性更為明顯(圖2,3,5)。這與其他研究一致,即資源可用性的時間波動解釋了為什麽微生物對時間變量的響應大於對空間變量的響應。研究地點微生物資源可用性的時間波動可能源於短生長季節植被生長的單峰模式。此外,本研究發現土壤濕度和質地對分解者多樣性-功能關系的影響是一致的(圖5)。濕度和質地可以在棲息地過濾中發揮重要作用,因為它們透過分類組成促進功能(圖5),導致分解者之間功能冗余的喪失。雖然濕度和質地的影響(圖5)與跨生態系的其他研究一致,但與其他研究不同,本研究沒有發現土壤pH值對這種關系的影響。這可能是由於采樣點(空間)土壤pH值的相對不變性,但不同景觀和生態系之間的差異可能更大。
高度多樣化的微生物群落結構和功能缺乏空間異質性會導致功能冗余。舉個例子,本研究平均每個樣本記錄了700多個OTUs,在760萬個reads中總共記錄了1937個OTUs。因此,可以理解的是,數據沒有顯示物種豐富度和功能之間的關系(圖S4)。例如,在每個樣本的局部豐富度低於200 OTUs的地方,有證據表明存在耦合,因此缺乏冗余。但是,在每個樣本700 OTUs的情況下(圖3-4),微生物多樣性可能與功能解耦。因此,毫不奇怪,不同的研究已經遇到了微生物物種豐富度和功能之間的耦合和解耦關系,本研究的數據強調了適應物種組成的重要性。
對於高度多樣化的自然微生物群落,冗余的存在經常受到質疑。從嚴格意義上講,功能冗余指的是,在一個群落中,存在執行重疊功能的生物體,它們可以在不改變功能的情況下很容易被取代。然而,在自然群落中很少出現這種情況,因為不同生物執行同一組功能的環境閾值(如溫度、降雨)可能不同。例如,本研究發現季節性和土壤因素(濕度和質地)對多樣性-功能關系有很強的控制作用。此外,功能冗余的概念還取決於研究了哪些功能,以及有多少功能(圖6)。一方面,在自然群落中,微生物可以同時執行多種功能,其中一些功能會被其他微生物共享,從而使功能冗余。另一方面,有些功能可能只針對少數幾個特定的物件,這使得功能特化且不冗余。因此,本研究的結果不是完全冗余,而是部份冗余,其中一些功能可能由自然群落中的多個微生物執行,而不是全部。但是,由於1937個OTUs尚未被解析到屬/種水平,我們並不完全了解負責高級功能的微生物的身份。然而,預期的技術進步可以彌補這種知識空白,從而更好地解釋這一關系。
鑒於土壤微生物分類的分辨率較低,以往的研究往往依賴於宏基因組的解釋和從各種蛋白質和酶的遺傳密碼的存在/缺失中推斷假定的功能。這並不能保證在樣本中實際執行相應的功能,因為「所有的東西都是無處不在的,但環境選擇不是」。本研究透過測量功能的大小來評估多樣性-功能關系,盡管只有30個與碳化合物分解代謝相關的功能中的一小部份。但我們的思維實驗表明,與宏基因組方法的103個假定功能相比,對101個功能的關系解釋是可推廣和可靠的。此外,微生物和分解的這種耦合的多樣性-功能關系與生產者和生產的模式一致,其中物種的身份比物種的數量更重要。
本研究發現微生物多樣性和功能之間的關系與大型哺乳食草動物的存在與否有關。這也符合生產者及其生產中的恢復力和抗性模式,即排除食草動物不會影響生產者的多樣性和生產。然而,與消費者對生產者的直接影響相反,分解者的運作機制可能不同。可能存在多種相互依賴的路徑來支持這種依賴性和抗性。首先,食草動物直接和間接影響的相對優勢之間的平衡可以表現為對分解者多樣性-功能關系沒有凈影響。其次,由於競爭-合作相互作用,微生物多樣性和功能之間的緊密耦合可能在我們十年之久的食草動物排除實驗開始之前就已經建立起來了。一旦建立,微生物分解者之間的相互作用可能經受住實驗性的食草動物排除(抗性),並/或從中迅速恢復(恢復力)。第三,食草動物對本研究中未量化的其他類別的分解者的影響可能不同,例如古菌和真核分解者。
總體而言,本研究發現微生物群落組成的差異會影響微生物功能多樣性,而不影響生物物種豐富度。這與功能冗余的一個關鍵方面不一致,即假設物種多樣性(豐富度和組成)與功能多樣性之間沒有關系。相反,本研究結果表明,微生物群落中某些物種(即組成)的減少或增加對微生物功能的影響比其他物種更大。因此,物種多樣性緩沖(即抗性)和改善(恢復力)壓力源效應的潛力取決於未受壓力源影響的物種的身份。雖然微生物多樣性-功能關系似乎對實驗性移除大型哺乳食草動物具有抗性和/或恢復力,但它們對土壤濕度的變化很敏感,而土壤濕度是大多數旱地生態系過程和服務功能的主要決定因素。未來的研究可以解決微生物功能的抗性和恢復力如何與全球變化的各個方面相吻合,包括降雨量的年際變化及其年內再分配。旱地降雨持續變化對土壤水分有效性的影響,特別是在跨喜馬拉雅和中亞的高海拔旱地,可能會對其生態系功能和服務的保護和可持續管理構成挑戰。











