文章資訊

摘要
北溫帶植物區系洲際間斷最顯著的例子之一涉及東亞和北美東部和西部。盡管150多年來人們對這種植物地理格局產生了相當大的興趣(例如,Gray,1859;Li,1952;Graham,1972;Boufford和Spongberg,1983;Wu,1983;Tiffney,1985a,1985b),但類群之間的關系顯示這種間斷仍然是模糊的。然而, 了解系統發育關系是對這種分布模式進行歷史生物地理學分析的先決條件。 為了更好地了解顯示這種洲際間斷的類群之間的關系,我們使用東亞、北美東部和西部的四個屬(山茱萸屬、八幡草屬、黃水枝屬和唐松毛茛屬)的各種DNA數據集進行了系統發育分析。為四個屬中的每一個都構建了面積分支圖,所有屬都顯示出相似的關系模式:東亞物種是所有北美物種的姐妹。在表現出這種間斷的其他三個類群(楤木屬、夏蠟梅屬和掌葉鐵線蕨)中也發現了相同的系統發育模式。在這七個不同的屬中發現的一致的系統發育模式提出了東亞、北美東部和西部間斷的共同起源的可能性。這些數據與長期以來的假設一致,即這種 著名的植物區系間斷代表了曾經連續的混合中生林群落的破碎化,並表明這種間斷可能只涉及兩個主要的間接事件 :歐亞大陸和北方之間的最初美洲分化,其次是北美東部和西部之間的植物區系隔離。然而, 系統發育和地理分布之間的一致性並不一定表明相同的植物地理歷史。表現出相同系統發育模式的類群可能起源於不同的地質時期。 利用分子鐘對分化時間進行分析表明,山茱萸屬、八幡草屬和夏蠟梅屬的物種可能在不同的地質時期發生了分化,這表明涉及 東亞和北美的區系間斷可能並不簡單;它可能涉及不同屬的不同地質時期的多個歷史事件。
引言
北溫帶的植物區系表現出驚人的洲際植物區系相似性。這種相似性在東亞和北美東部之間最為顯著, 在以下五個地區中的兩個或多個地區也有較小程度的相似性:東亞、西亞、北美東部、北美西部和東南歐。北溫帶表現出洲際間斷的植物區系元素通常被認為是第三紀遺跡,在中新世晚期之前更廣泛地分布在整個勞亞大陸,形成混合中生林群落的一部份(Li,1952;Wolfe和Leopold,1967;Wolfe,1969年、1972年、1975年;Wood,1971年、1972年;Graham,1972年。根據這一假說,分裂是隨著地理和氣候變遷而出現的,包括北美和歐亞大陸因大陸漂移而間斷、漸新世舊世界圖爾蓋海峽的關閉、北美西部的第三紀造山運動以及全世界的造山運動。第三紀的氣候波動。這些事件消除了許多地區的植物,特別是歐洲和北美中部(Graham,1972;Leopold和MacGinitie,1972;Tiffney,1985a)。
北溫帶植物區系的間斷分布模式,特別是那些涉及東亞和北美的分布模式,150多年來一直是研究的主題(例如,Gray,1859、1878;Hu,1935;Chaney,1947;Li,1952,1972;Graham,1972;Wolfe,1972,1981,1983;Iltis,1983,1993;1983年;Koyama,1983;Tiffney,1985a,1985b;)盡管人們長期以來一直感興趣,但顯示間斷的類群之間的系統發育關系仍然模糊,並且間斷的起源一直存在爭議。在以下方面存在主要分歧: (1)表現出間斷的植物之間的關系模式(如果有),(2)間斷是否具有單一起源或涉及不同分類群中不同時間的多個生物地理事件,以及(3)擬議的間斷類群的地理起源。 在正確評估有關這種突出的生物地理間斷的這些問題和其他問題之前,需要清楚地了解表現出間斷模式的類群的系統發育。
我們關註的是在東亞和北美東部和西部顯示出不連續分布的類群。這三個地區大約有30個屬具有密切相關的物種(Wood,1971、1972;Li,1972;Wu,1983)。對於在這三個地區發現的植物如何相互關聯,目前還沒有達成共識;作者為不同的分類單元提出了不同的模式。Tamura(1983)在唐松毛茛屬中指出,最接近的親緣關系是東亞物種和北美西部物種之間。相比之下,在水甘草屬、白珠屬、香根芹屬和安息香屬中,Wood(1972)認為東亞和北美東部份類單元之間關系最密切。這些假設僅基於形態學相似性,沒有進行系統發育分析。
僅對亞洲東部和北美東部和西部的少數屬進行了系統發育研究,包括嗩吶草屬(Soltis等人,1991;Soltis和Kuzoff,1995)、繡球屬(Soltis等人,1995)、掌葉鐵線蕨(Paris,1991;Paris和Haufler,1994)、延齡草屬(Kato等人,1995)、夏蠟梅屬(Wen等人,1996)和楤木屬楤木(五加科)(Wen等,1996)。系統發育研究表明, 長期以來被認為是這種植物區系間斷的「經典」例子的繡球屬和嗩吶草屬是多系的。 因此,這些屬並不是研究這種析取的有用模型。這些發現清楚地證明了健全的系統發育框架在推斷生物地理歷史中的重要性;對並非真正近親的類群進行比較可能會導致錯誤的生物地理學結論。相比之下,延齡草的系統發育分析表明,盡管是單系的,但植物地理學假說最好在該屬的離散分支內考慮(Kato等人,1995),這項任務需要比目前可用的更大的分類單元密度。分析夏蠟梅屬(三種,每個區域一種)和楤木屬(九種,一種在北美東部,一種在北美西部,七種在東亞)表明來自北美東部和西部的物種是姐妹,而這些物種又是亞洲物種的姐妹。顯然, 需要對更多顯示這種間斷分布模式的類群進行系統發育研究,以提高我們對東亞、北美東部和西部間斷的理解。 因此,我們對四個屬進行了分子系統發育分析,即山茱萸屬(山茱萸科)、八幡草屬和黃水枝屬(虎耳草科)以及唐松毛茛屬(毛茛科)的大苞山茱萸屬,這些屬目前僅出現在亞洲東部、北美東部和西部。我們的目標是:(1)評估這四個屬的系統發育,(2)確定不同屬是否存在顯示這種分布模式的一般系統發育模式;(3)根據這些屬的分子系統發育構建區域分支圖,以闡明東亞、北美東部和北美西部之間的關系;(4)初步了解表現出這種間斷的類群的地理起源,以提高我們對北溫帶植物區系的了解。
材料與方法
分子系統發育分析
對幾個DNA區域進行了測序以推斷系統發育,但並非每個屬的所有區域都進行了分析:葉綠體基因rbcL和matK以及核糖體RNA基因的內部轉錄間隔區(ITS區域)。擴增和測序方法遵循Morgan和Soltis(1993)以及Xiang等人的方法。(1993)對於rbcL,Johnson和Soltis(1994,1995)對於matK,以及Baldwin(1992)和Soltis和Kuzoff(1995)對於ITS。先前發表的cpDNA限制性位點變異也用於系統發育評估(Soltis等,1991、1993;Xiang等,1996)。
除非圖例中特別說明,系統發育分析的一般方法如下。對於每個屬,使用了幾個外群。此外,還進行了涉及相關屬的廣泛系統發育分析和僅涉及研究屬的更集中的分析。狹義分析的外群是根據確定研究組姐妹類群的廣義分析的結果選擇的。在所有分析中,每個屬內的系統發育關系模式都是相同的。
使用Fitch簡約法透過PAUP3.1.1(Swofford,1993)對數據進行分析。對於山茱萸屬和八幡草屬,進行了分支定界搜尋,對於黃水枝屬和唐松毛茛屬,使用了窮舉搜尋選項。為了評估最簡約樹中揭示的關系的相對支持度,進行了100個重復的引導分析(Felsenstein,1985),並根據Eer-nisse和Kluge(1993年)評估了Bremer支持(或衰減指數)(Bremer,1988))。下面列出了所分析的每個屬的其他背景數據。
山茱萸。 山茱萸(山茱萸)是一個大約有55個物種的屬,其中大苞山茱萸形成一個單系類群(Xiang等,1993,1996;Xiang和Soltis,出版中)。該類群由七個密切相關但形態各異的物種組成,其中多花狗木分布於北美東部,盤狀花梾木分布於中美洲,太平洋狗木位於北美西部,以及頭狀四照花、四照花、香港四照花和多脈四照花位於東亞(Xiang,1987)。大苞山茱萸物種之間的形態變異與其地理分布相符。所有東亞物種在形態上都非常相似,並在頭部產生復合果實。在最近使用形態特征的系統發育分析中,這些物種也形成了一個獨特的分支(Murrell,1993)。所有美洲物種都成簇地結出單獨的果實。來自北美東部和東亞的山茱萸有四個大而艷麗的苞片,而來自北美西部的太平洋山茱萸有六個大而艷麗的苞片;來自中美洲的盤狀四照花有四個苞片,早期脫落但未擴充套件。
系統發育分析采用cpDNA限制性位點和rbcL-matK序列cpDNA限制性位點的組合數據集(1440bp的rbcL、1212bp的matK和242個限制性位點)。所有三個美洲物種,多花狗木、盤狀四照花和太平洋山茱萸,以及東亞支系的兩個代表頭狀四照花和四照花,均包含在cpDNA限制性位點分析中。然後,除盤狀四照花之外的所有這些物種(由於早期限制性位點研究後DNA降解,rbcL和matK無法擴增,因此未包括在內),然後將所有這些物種納入rbcLmatK序列和cpDNA限制性位點的聯合分析中。組合數據集分析中包含的所有分類單元至少具有三個可用數據集中的兩個。根據以前對山茱萸及其近親的研究結果(Xiang等,1996,1998a),兩種矮山茱萸(草茱萸和矮山茱萸)和三種山茱萸(歐洲山茱萸、常見山茱萸和山茱萸屬sessilis)被選為外群。分子數據來自我們最近的研究(Xiang等人,1993,對於rbcL序列;1996,對於限制性位點;以及1998a,對於matK序列),除了本研究中生成的山茱萸rbcL序列(表1)。
八幡草屬。 八幡草屬包含7個物種,其中包括八幡草屬aconitifoliaNutt。來自北美東部的唯一物種八幡草東亞的單一種,以及五個種:八幡草屬intermedia(Piper)G.N.Jones、八幡草屬MajorA.Gray、八幡草屬occidentalis Torrey&Gray、八幡草屬rotundifoliaParry和八幡草屬richardsonii(Hook.)Rothrock,北美西部(Gornall和Bohm,1985;Soltis等,1993)。八幡草屬的單系性已在先前對虎耳草科(Saxifragaceae)的幾次系統發育分析中得到證實。(Soltis等,1993、1996;Johnson和Soltis,1995)。我們重點關註屬內的關系,並分別使用cpDNA限制位點和核ITS序列進行系統發育分析。除八幡草屬richardsonii外,該屬的所有物種均包括在內,該種高多倍體尚無合適的DNA(參見Soltis等,1993)。葉綠體DNA限制位點數據來自Soltis等人(1993),ITS序列來自Soltis等人(1996)。根據先前對虎耳草科的分析結果,使用堇蓬草屬和洪崖草屬作為外類群。(Soltis等人,1993)。由於cpDNA和ITS樹揭示了北美分類群之間的不同關系,因此這兩個數據集並未合並為單一分析。
表1 本研究中用於系統發育分析的材料和分子數據的來源
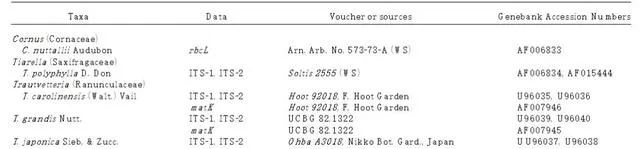
註.UCBG、Arn的縮寫。Arb.和RBG分別代表加州大學植物園、阿諾植物園和英國邱園皇家植物園。Xiang和Soltis代金券存入WS。
黃水枝屬。 黃水枝屬有3個種:北美洲東部的黃水枝屬cordifolia L.,北美洲東部的黃水枝屬trifoliata L.,有3個變種(var.trifoliata、var.unifoliata(Hook.)Kurtz.和var.laciniata(Hook.)Wheel.)。北美洲西部和亞洲東部的黃水枝屬polyphyllaD.Don。先前對幾種葉綠體標記的分析表明黃水枝屬是多系的(Soltis等,1991;Xiang,1995)。這些分析表明,北美東部物種(黃水枝屬cordifolia)與礬根屬的關系比與其他黃水枝屬物種的關系更為密切。相反,形態學和核ITS序列數據表明黃水枝屬是單系的(Soltis和Kuzoff,1995)。全部數據表明,黃水枝屬cordifolia顯然捕獲了礬根屬物種的葉綠體,並且基於cpDNA的拓撲不能反映生物體關系。正如其他地方所評論的那樣(Soltis和Kuzoff,1995),黃水枝屬cordifolia和礬根屬物種之間自然發生的屬間雜種的存在進一步表明了這種葉綠體捕獲情況。
由於葉綠體基因組對黃水枝屬中的有機關系的評估不準確,因此我們的系統發育假設僅基於核序列。在這項研究中,我們使用ITS序列對黃水枝屬的三個物種進行了系統發育分析。首先使用雨傘草屬和鬼燈檠屬作為外群,對整個礬根屬(由黃水枝屬和虎耳草科其他八個屬組成的分支;Soltis等人,1991,1993)進行了廣泛的ITS序列分析。ITS序列來自Soltis和Kuzoff(1995),除了本研究中獲得的黃水枝的序列之外。因為我們對ITS序列的廣泛分析表明黃水枝屬是單系的(樹未顯示),所以使用詳盡的搜尋對黃水枝屬進行了第二次狹義的ITS序列分析。崖葵草、異葉嗩吶草和裸米嗩吶草被用作本次重點分析的外類群,因為它們與黃水枝屬具有密切的系統發育關系。
唐松毛茛屬。 唐松毛茛屬由三個物種組成:來自北美東部的唐松毛茛屬carolinensis(Walt.)Vail、T.grandisNutt。來自北美西部和唐松毛茛屬japonica Sieb. & Zucc。來自東亞。獲得了這三個物種以及外群鼠尾毛茛的matK和ITS序列。基於對毛茛科的廣泛系統發育分析結果(Hoot,1995,個人通訊),選擇鼠尾毛茛作為外群,表明鼠尾毛茛是唐松毛茛屬的近親。Johnson和Soltis(1994,1995)為matK設計的測序引物中,只有1168R和1470R兩種可以用於本研究。這些引物為所研究的物種提供了大約600個堿基對(bp)的matK序列。還獲得了唐松毛茛屬和鼠尾毛茛的ITS-1和ITS-2區域的序列。對matK和ITS序列進行了單獨分析。由於ITS和matK序列產生相同的拓撲結構,因此我們組合了這些數據集並對組合的數據矩陣進行了後續分析。
面積分支圖
透過用相應分類單元的分布面積替換分支圖上的末端分類單元名稱,為山茱萸、八幡草屬、黃水枝屬和唐松毛茛屬的大苞山茱萸構建了區域分支圖(參見Platnick和Nelson,1978;Nelson和Platnick,1981;Humphries和Parenti,1986)。比較了四個屬的面積分支圖,並構建了一致的一般面積分支圖,以顯示東亞、北美東部和北美西部之間的面積關系。
起源中心
在沒有系統發育資訊的情況下建立一個群體的起源中心(例如,使用「年齡-區域」假設;參見Brown和Gibson,1983;Futuyma,1986)通常是有問題的。我們嘗試使用MacClade3.05(Maddison和Maddison,1992)的系統發生方法來做到這一點,以最簡約的方式將字元「區域」對映到拓撲上,以推斷祖先(根節點)的地理區域每個學習小組。然而,在許多情況下,進行這種繪圖工作非常簡單,可以手動完成。
由廣泛的系統發育分析產生的拓撲清楚地描繪了研究組的姐妹組(這些樹此處未顯示)被用作確定「區域」對映拓撲中使用的外群的基礎。對於山茱萸,透過對整個屬的cpDNA限制性位點進行廣泛分析而得到的拓撲結構以及代表整個山茱萸科的組合rbcL-matK數據集用於確定大苞山茱萸的姐妹群(Xiang等,1993,1998a)。對於八幡草屬,整個「八幡草屬群」(六個屬的前進演化枝)的ITS和cpDNA限制性位點拓撲(Soltis等人,1993,1996),以及代表整個「八幡草屬群」的cpDNA限制性位點和matK拓撲虎耳草科s.s。(Soltis等人,1993年,1996年)。對於黃水枝屬,我們使用了整個礬根屬(九個屬的分支)的ITS樹(Soltis和Kuzoff,1995)。對於唐松毛茛屬,我們使用了毛茛科的組合atpB-rbcL18SrDNA樹(Hoot,1995)。外群的地理分布是根據外群所有成員的已知分布(而不僅僅是圖中描繪的幾個外群物種)來分配的。因此,系統發育樹中顯示的物種的地理區域不一定代表整個外群團的地理區域。例如,透過對該屬的廣泛分析,將大苞山茱萸的姐妹群確定為矮山茱萸。矮山茱萸有四種分布在環流地區並延伸到緬甸(參見Xiang等,1996)。在系統發育樹(圖1)中,僅顯示了兩個北美物種:草茱萸和矮山茱萸。在地理「區域測繪」分析中,大苞山茱萸外群的地理區域被認為是環北陸界,而不是北美洲。
我們也對金盞花和楤木進行了同樣的練習。楤木,由其他人進行系統發育研究的分類單元(Wen等人,1996)。對於夏蠟梅屬,采用了代表木蘭科植物的rbcL序列的廣泛系統發育分析(Qiu等人,1993)。對於楤木屬,該科的cpDNA限制性位點拓撲結構,采用楤木屬Dimorphanthus作為姐妹群(Wen等人,1996)。
結果
系統發育關系
山茱萸。 對大苞山茱萸cpDNA限制位點的系統發育分析揭示了與整個山茱萸屬廣泛分析中發現的關系相同的關系(圖1a)(Xiang等,1996)。也就是說,取樣的東亞物種(頭狀四照花、四照花)形成一個前進演化枝,並且是包含所有美洲物種的前進演化枝的姐妹群。對大苞山茱萸的組合rbcL-matK-cpDNA限制性位點數據集的分析揭示了僅用限制性位點觀察到的相同的一般拓撲結構(盡管現在省略了盤狀四照花;見上文)(圖1b)。東亞物種(頭狀四照花、四照花)形成一個分支,是所分析的北美物種多花狗木和太平洋山茱萸的姐妹。
八幡草屬。 八幡草屬的cpDNA限制性位點和ITS序列的系統發育分析均將東亞物種八幡草屬lycoctonifolia視為包含所有北美物種的前進演化支的姐妹支(圖2a和2b)。在這個北美分支中,ITS樹將來自北美西部的八幡草屬rotundifolia視為包含來自北美東部的八幡草屬aconitifolia和其余北美西部物種的分支的姐妹(圖2b),而cpDNA樹顯示八幡草屬aconitifolia、八幡草屬rotundifolia和其余物種之間的三分法(圖2a)。
黃水枝屬和唐松毛茛屬。 黃水枝屬的ITS序列的系統發育分析以及唐松毛茛屬的ITS和matK序列分析表明,對於這兩個屬,這兩個北美物種彼此之間的相關性比其中一個與東亞物種的相關性更密切(圖3和圖4))。
面積分支圖
基於大苞片物種山茱萸屬、黃水枝屬、八幡草屬和唐松毛茛屬的分子系統發育的四個區域分支圖是一致的,表明北美東部和西部彼此之間的關系比東亞和東亞之間的關系更為密切(圖5)。
起源中心
我們嘗試透過將地理區域對映到拓撲上來推斷這四個屬的起源中心,已經產生了一些結果,這些結果在很大程度上與研究組姐妹組的地理分布相關(見圖1-4)。例如,八幡草屬以及虎耳草科的所有廣泛系統發育分析。(Soltis等人,1993、1995、1996)表明,八幡草屬的姐妹類群是洪崖草屬和堇蓬草屬,這兩個屬僅限於北美西部。因此,對於八幡草屬來說,區域測繪表明其起源中心是北美西部。相比之下,在山茱萸中,大苞山茱萸的直接姐妹群是矮山茱萸,這是一種環北分布的類群。因此,大苞山茱萸的原產地是模棱兩可的。同樣,對於唐松毛茛屬來說,其姐妹屬鼠尾毛茛和毛茛屬(Hoot,1995)都廣泛分布在北半球。因此,該屬的地理起源再次被表明是不確定的。對於黃水枝屬,尚未確定得到良好支持的姐妹團體(Soltis和Kuzoff,1995)。尋找黃水枝屬的姐妹群是很復雜的, 因為在其屬的前進演化枝中大量葉綠體捕獲使得cpDNA數據對於系統發育推斷毫無用處,並且ITS序列數據無法充分解析黃水枝屬的關系。因此,該屬的起源中心也仍然不確定。
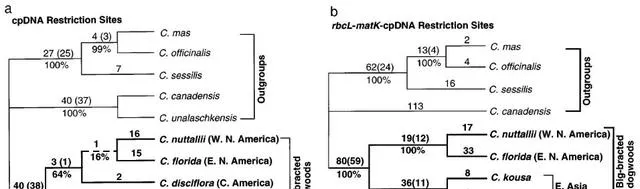
如圖1. 山茱萸大苞山茱萸的系統發育分析。(a)使用山茱萸(歐洲山茱萸、山茱萸和茱萸屬sessilis)和矮山茱萸(草茱萸和茱萸屬unalaschkensisas)對cpDNA限制性位點數據進行詳盡搜尋後得到的三棵最矮的樹之一外群(長度5175個步驟;CI50.961,排除無資訊字元,RI50.982)。內組以粗體指定。每個分支上方給出了支持每個前進演化枝的限制性位點突變的數量,以及衰減值(在括弧中);引導值在下面的分支中給出。虛線代表所有三棵最短的樹中未辨識的分支。(b)對大苞山茱萸的rbcL-matK序列和cpDNA限制性位點進行聯合分析得出的單棵最短樹(長度5405步;CI50.938,排除無資訊性狀,RI50.962)。內組以粗體指定。分支機構的編號如(a)所示。
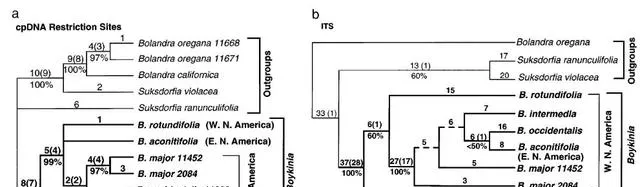
如圖2. 八幡草屬的系統發育分析。(a)八幡草屬cpDNA限制性位點的系統發育分析產生的單個最短樹(長度557個步驟;CI50.911,排除無資訊性狀,RI50.965)。內組以粗體指定。分支上的編號如圖1a所示。(b)八幡草屬ITS序列系統發育分析產生的兩棵最短的樹之一(長度5236步;CI50.747,排除無資訊性狀,RI50.802)。內組以粗體指定。分支上的編號如圖1a所示。虛線表示在兩棵最短的樹中都無法辨識的分支;所有其他分支都存在於這兩棵最節儉的樹中。
對於夏蠟梅屬來說,姐妹類群是蠟梅屬(蠟梅科唯一的其他成員),該屬僅限於東亞(Qiu等人,1993)。因此,亞洲被認為是金盞花的起源中心,楤木屬的姐妹團體。楤木屬Dimorphanthus(Wen等人,1996),分布於亞洲東部和北美東部。盡管根據其姐妹群的分布,楤木屬的起源地點是模棱兩可的,但楤木屬內基礎譜系的分布表明其起源於亞洲。楤木屬的第一個分支終止於來自東亞的物種。
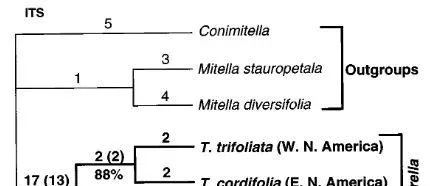
如圖3. 黃水枝屬ITS序列系統發育分析得出的單個最短樹(長度539步驟,CI50.952,排除無資訊性狀,RI50.971)。內組以粗體指定。分支上的編號如圖1a所示。
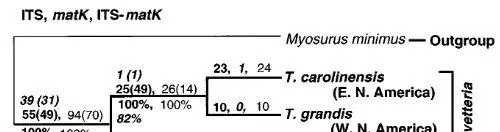
如圖4. 分別對唐松毛茛屬的ITS、matK和組合ITS-matK序列進行系統發育分析,得出相同的最短樹。單棵ITS最短樹的長度為120步,CI為0.974(排除無資訊字元),RI為0.971;單個matK最短樹的長度為46步,CI為1.000(排除無資訊字元),RI為1.000;單個ITS-matK樹的長度為166個步驟,CI為0.983(不包括無資訊字元),RI為0.982。內組以粗體指定。分支上的編號如圖1a所示;斜體字代表matK樹,羅馬字代表ITS-matK。
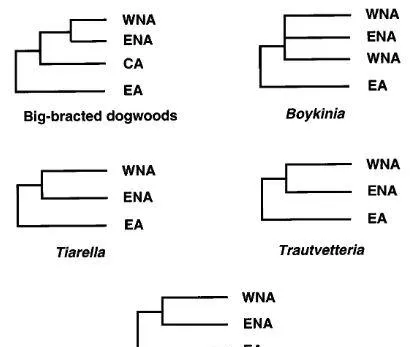
如圖5. 透過將系統發育樹上的末端分類單元名稱替換為相應分類單元的分布區域而獲得面積分支圖。WNA,北美西部;ENA,北美東部;CA,中美洲;和EA,東亞。
Michael J.Donoghue院士正在講解該篇文章
討論
關系模式。 對亞洲東部、北美東部和北美西部間斷出現的四個屬(山茱萸屬、八幡草屬、黃水枝屬和唐松毛茛屬的大苞種)的系統發育分析揭示了物種之間生物地理關系的單一模式(圖1-4))。在所有四個屬中,北美東部的物種是北美西部物種的姐妹群,亞洲物種是所有北美物種的姐妹群。相反,基於其他數據來源的直觀推論(即非系統發育)表明存在替代關系,或者沒有解決每個屬內的關系。例如,黃水枝屬的三個物種具有非常相似的核型,並且各自具有一組獨特的類黃酮成分(Soltis和Bohm,1984)。這些特征沒有在黃水枝屬中提供任何共源性,並表明這三個物種同樣不同。基於形態特征對唐松毛茛屬關系的非分支推斷表明,東亞物種(日本黃水枝)和北美西部物種(黃水枝屬grandis)彼此之間的親緣關系比其中任何一個物種與北美東部物種(黃水枝屬grandis)之間的親緣關系更為密切(黃水枝屬carolinensis)(Tamura,1983)。這一結論與此處提出的系統發育分析相沖突。
這裏報道的四個屬的系統發育關系模式與楤木屬相似。楤木屬(Wen等人,1996)、夏蠟梅屬 (Wen等人,1996)和掌葉鐵線蕨(鐵線蕨屬)(Paris,1991;Paris和Haufler,1994),均具有物種或分布於東亞、北美東部和北美西部的族群。因此,系統發育分析表明,山茱萸屬、八幡草屬、黃水枝屬、唐松毛茛屬、楤木、夏蠟梅屬和掌葉鐵線蕨等大苞物種,這些不同的類群,都表現出相似的物種間關系模式: 北美東部和西部物種始終是姐妹,亞洲代表又是北美分支的姐妹。
目前還不清楚這裏觀察到的系統發育模式在表現出這種間斷的大約30個屬中可能有多普遍。因此,鼓勵對表現出這種間斷的類群進行額外的系統發育分析。還值得註意的是, 大量屬表現出比此處討論的稍微復雜的析取模式。也就是說,除了在東亞和北美東部和西部有代表外,一些屬在歐洲也有成員分類單元 (例如七葉屬、金腰屬)。盡管如此,最近的一些系統發育分析表明,七葉樹屬(七葉樹科;ITS和matK序列數據;Xiang等人,1998b)和金腰屬(HibschJetter等人,未發表)可能存在與此處提到的相似的系統發育模式。因為北美物種形成了一個與舊世界分類單元姐妹的前進演化枝。
一致的生物地理模式與共同的植物地理歷史。系統發育拓撲和譜系地理分布之間的一致性已被用作共享歷史生物地理學推斷的基礎(Platnick和Nelson,1978;Nelson和Platnick,1981;Humphries和Parenti,1986;Sober,1988;Oosterbroek和Arntzen,1992)。基於這一原理,在此處比較的七個不同類群中觀察到的一致的系統發育關系模式可能表明共同的生物地理歷史。例如,這些數據可以被視為對長期存在的假設的支持, 即東亞、北美東部和北美西部的間斷代表了曾經連續的植物群落的分裂 (例如,Gray,1878;Hu,1935;Chaney,1947,1959;Wood,1971,1972)。例如,Gray(1878)提出,穿過白令海峽的北半球高緯度地區存在著連續的植物群。該植物群隨後被更新世冰川作用破壞,導致不同大陸上的類群分裂。Gray的假說由Chaney(1947,1959)進一步發展,他提出了「北極-第三紀地植物區系」的概念來解釋東亞和北美之間的植物區系相似性。錢尼設想了一個常見的生物群,其分類組成與現代北溫帶植物群相似,占據了第三紀早期歐亞大陸和北美的高緯度地區。這種地理植物群隨後向南傳播,其組成幾乎沒有變化。 在第三紀晚期和第四紀期間,歐洲、西亞和北美中部的大部份植物群滅絕,導致今天在北半球觀察到的間斷。 然而,最新的古生物學證據與北極-第三紀地植物區系概念相矛盾(Wolfe,1969、1972、1975、1985)。新的古生物學證據表明,北半球存在具有動態分類組成的第三紀早期「向水植物群」(Wolfe,1969、1972、1975、1985)。Wolfe(1969,1972,1975,1985;另見Tiffney,1985a,1985b)認為北溫帶植物區系的洲際相似性是向溫帶植物區系擴散的結果。這種向北植物區系在第三紀中期發展成為所謂的「混合中生林」(一種暖溫帶植被類別)(Wang,1961;Tiffney,1985a);這片森林曾經或多或少連續分布在整個北半球,但後來的氣候和地質變化導致其範圍受到限制,並最終導致今天觀察到的間斷(Li,1952;Wood,1971,1972;Graham,1972;Wolfe,1972,1981;Hsu,1983;Tiffney,1985a,1985b)。因此,現存的東亞-北美東部和西部的斷層被認為是這種混合中生林的殘余部份(Li,1952;Wood,1971、1972;Graham,1972;Hsu,1983)。根據這一假設,觀察到的系統發育關系模式表明,北美東部和西部的間斷發生在東亞-北美最初的隔離之後。因此, 連續的中生混合森林的最初破壞之一就是由於大陸漂移而使歐亞大陸與北美間斷 。後來各種地質和氣候變遷使這種植物群從歐洲大部份地區、中亞和北美中部消失(Graham,1972;Leopold和MacGinitie,1972;Tiffney,1985a,1985b),導致東亞、東部和西部的間斷。北美。
然而,我們強調,系統發育和地理分布之間的一致性並不一定表明相同的植物地理歷史。 亞洲東部、北美東部和北美西部的植物區系間斷可能起源於不同屬的非常不同的地質時期,這種現象被稱為「偽一致性」( Cunningham和Collins,1994),Tiffney指出(1985a;另見Wolfe,1969、1972、1981;Li,1972)。在這方面,關於不同屬物種之間分化時間的資訊對於確定在不同的七個類群中觀察到的 相同系統發育模式是否代表假一致性或共享的植物區系歷史至關重要。
譜系之間的分歧時間可以使用化石證據或分子鐘來評估(盡管分子鐘方法的困難是眾所周知的;見下文)。本文討論的所有七個類群,除了山茱萸之外,要麽是未知的化石(八幡草屬、黃水枝屬、唐松毛茛屬和掌葉鐵線蕨),要麽沒有足夠的化石記錄(楤木屬和夏蠟梅屬)來評估不同物種之間的分歧時間(Collinson等人,2017,1993;Taylor和Taylor,1993;Friis等,1994;Mai,1994)。歐洲漸新世中期和中新世沈積物中發現了單果大苞山茱萸(如現存的北美物種)的果核化石,並行現了復果大苞山茱萸(如現存的東亞物種)的果核歐洲的上新世和日本的更新世(參見Eyde,1988)。這些化石證據表明,大苞山茱萸在距今(BP)大約30-3200萬年的漸新世期間前進演化而來。化石證據還表明,東亞和北美的大苞山茱萸距今至少有五百萬年的分化。
分子鐘基於分子前進演化的中性理論,並假設不同譜系的分子前進演化速率恒定(Zuckerkandle和Pauling,1965;Kimura,1983)。盡管關於中性理論存在爭議,並且不同譜系中分子前進演化的異質速率已被記錄(例如,Wilson等人,1990;Gaut等人,1992、1993、1996),但分子鐘可能有助於如果可以有一定信心地校準時鐘,則可以評估發散時間。據評估,葉綠體基因組每年每個位點的總體分化率約為1029個核苷酸取代(Zurawski等人,1984年;Zurawski和Clegg,1987年)。該分子鐘用於根據cpDNA限制性位點數據評估鵝掌楸屬(木蘭科)東亞-北美東部間斷物種對之間的分化時間(Parks和Wendel,1990)。該評估與化石證據(距今11-1400萬年)的評估一致(Parks和Wendel,1990)。
為了深入了解東亞和北美東部和西部物種之間的分化時間,我們將這種分子鐘套用於三個屬:山茱萸屬、八幡草屬和花萼屬,這些屬都有cpDNA限制位點數據。假設序列分歧率為每百萬年0.1%,根據2.61%的序列分歧評估,東亞和北美大苞山茱萸之間的分歧時間評估為1310萬年BP(中中新世)使用電腦程式SDE1.2(Wolfe和Wolfe,1993)從限制性位點突變中獲得[SDE程式使用Jukes和Cantor(1969)方法評估序列分歧,並對多次命中進行校正。計算中采用了Nei(1987)的方程式5.3、5.38和5.41,以及Li和Graur(1991)的方程式3.19、3.32和3.33。]這一評估落在基於所討論的山茱萸化石證據評估的時間範圍內。多於。根據物種間平均1.94%的序列分歧,北美大陸大苞山茱萸的分化時間評估為距今970萬年(中新世中期)。
使用相同的分子鐘和cpDNA限制位點數據,東亞和北美八幡草屬物種之間的分歧時間評估僅為260萬年BP(上新世末期)(兩個譜系之間的序列分歧為0.51%);北美東部和西部物種之間的分歧時間為120萬年BP(更新世)(序列分歧為0.23%)。同樣,對於夏蠟梅屬,東亞和北美物種之間的分歧時間評估為距今310萬年,在上新世(0.61%序列分歧),而北美東部和西部物種之間的分歧時間為260萬年BP,或上新世末期(0.52%序列分歧)(Wen等,1996)。唐松毛茛屬、黃水枝屬和楤木屬的分歧時間。無法使用同一時鐘來評估楤木屬,因為涉及黃水枝屬和礬根屬的cpDNA捕獲的發生會扭曲評估,並且無法獲得唐松毛茛屬和楤木屬的cpDNA限制位點數據。
對分化時間的分析表明,山茱萸屬物種的分化早於八幡草屬和夏蠟梅屬物種的分化,而八幡草屬物種的分化最近。對 這些屬的物種的不同分歧時間的評估可能表明這些屬中的物種的間斷發生在不同的地質時期,或者,這些屬只是經歷了不同的分子前進演化速率。
為了區分這兩種可能性,需要對山茱萸屬、八幡草屬和花萼屬進行cpDNA前進演化的相對速率測試。如果對山茱萸屬、八幡草屬和夏蠟梅屬評估的不同分歧時間完全歸因於速率差異,那麽我們預計山茱萸屬的速率要高得多,但八幡草屬和夏蠟梅屬的速率相似。由於評估遠緣相關分類群中限制性位點突變的同源性變得有問題,因此對於屬於開花植物的三個不同亞類(分別為菊科、薔薇科和木蘭科)的這三個屬的限制性位點數據,此類測試是不可行的。因此,我們對這些屬的葉綠體基因rbcL進行了相對速率測試。使用Li和Bousquet(1992)的方法並遵循Gaut等人的方法,對同義和非同義替換進行了測試。(1996)和Xiang等人。(1998a)。Chase等人的rbcL序列分析中,金魚藻屬的rbcL序列是所有其他開花植物的姐妹屬。(1993),首先被用作測試的參考序列。因為金魚藻在Chase等人的rbcL樹中具有相對較長的分支(44)。(1993),它與所有其他開花植物的姐妹關系可能值得懷疑。因此,我們使用第二個參考序列,睡蓮的rbcL進行測試,睡蓮是Chase等人的rbcL分析中「PaleoherbII」的成員。(1993),比較結果。
當金魚藻屬作為參考分類單元時,無論是同義還是非同義替換,山茱萸屬和八幡草屬之間的rbcL前進演化率都沒有顯著差異(表2),但夏蠟梅屬和八幡草屬之間的同義和非同義替換則存在顯著差異(表2)。夏蠟梅屬的同義替換率低於八幡草屬,但非同義替換率則相反(表2)。山茱萸屬和金盞花屬也存在類似情況(見表2)。與夏蠟梅屬相比,山茱萸具有較高的同義替代率,但非同義替代率較低,這表明這些屬之間的總替代率可能沒有顯著差異。
表2 山茱萸屬、八幡草屬和蠟梅屬在rbcL下的相對速率測試結果
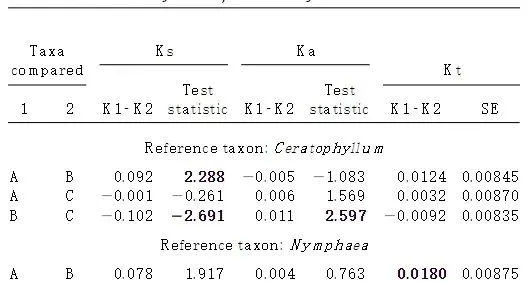
註.金魚藻和睡蓮被用作測試的參考類群。A、加拿大山茱萸;B、五花蠟梅;C,八幡草屬rotundifolia。測試中使用的類群的rbcL序列來自Xiang等人。(1993)為山茱萸屬,來自Chase等人。(1993)對於金魚藻屬、睡蓮屬和花蕊屬,來自Soltis等人。(1993)為八幡草屬。K1–K2,譜系1和譜系2之間每個位點的加權取代數差異;Ks,同義替換;Ka,非同義替換;和Kt,總換人數。對於Ks和Ka,絕對值為0.1.96(以粗體表示)的檢驗統計量在0.05水平上顯著。對於Kt,大於標準誤差兩倍的K1–K2值在0.05水平上被認為是顯著的。
我們隨後使用MEGA1.0的Kimura-2參數距離方法(Kumar等人,1993年)對總替代率進行的測試,繼Li(1997年,第218-219頁)之後,確實顯示三個屬之間沒有顯著差異(表2)。當睡蓮屬作為參考分類單元時,除了山茱萸屬和八幡草屬之間的非同義替代率存在顯著差異外,三個屬之間的同義和非同義替代率均沒有顯著差異(表2)。這些結果顯然不符合這樣的預期:山茱萸屬的較長分歧評估是由於rbcL前進演化速率升高所致。相反,為山茱萸屬、八幡草屬和夏蠟梅屬獲得的不同分歧時間可能為偽同余提供證據。也就是說,這三個屬在不同的地質時期在亞洲東部、北美東部和北美西部獲得了斷續分布。
因此,本研究中檢查的七個不同屬不一定具有相同的生物地理歷史,盡管它們在物種之間表現出相同的系統發育關系模式。一些屬,例如大苞山茱萸,可能是中生混合森林的遺跡,並且由於該物種在中新世中期附近分化,曾經連續分布在北半球。大苞山茱萸可能透過白令陸橋或北大西洋的一系列島嶼「墊腳石」分布在兩大洲(參見Tiffney,1985a,1985b)。其他物種,例如夏蠟梅屬(產生封閉在肉質容器中的種子,可能會被鳥類吃掉)和八幡草屬(產生可能被鳥類羽毛捕獲的結核種子),考慮到該物種可能透過長距離傳播獲得了不連續的分布。在這些屬中,最近根據分子鐘被分離出來。或者, 這兩個屬可能是透過逐漸遷移越過白令陸橋,然後在北美進行長距離擴散而獲得了間斷分布。白令陸橋幾乎在整個第三紀期間定期用於東亞和北美西部之間的植物交換 (直到距今350萬年;參見Allen,1983;Tiffney,1985a,1985b;Cunningham和Collins,1994)。相比之下,北大西洋大橋直到始新世晚期才發揮作用,盡管北大西洋中直到漸新世早期才存在一系列島嶼「墊腳石」(Tiffney,1985a)。北美東部和西部植物區系之間的元素交換在中新世停止(Tiffney,1985a)。因此,我們的分子數據並沒有反駁這樣的觀點,即東亞和北美之間的植物區系相似性具有復雜的歷史,涉及不同分類群中不同地質時期的多個歷史事件(替代和擴散)(Wolfe,1969,1972,1975,1981),1985;Li,1972;Tiffney,1985a)。
起源中心。 盡管我們已經在東亞和北美東部和西部發現的六個開花植物屬和一種蕨類植物之間發現了一致的系統發育關系模式,但我們的分析表明這些屬的起源大陸可能不同或根本不明確。我們透過強調基於地理分布到系統發育的對映來推斷起源中心的變幻莫測和困難來作為本次討論的序言。為金盞花和楤木屬。楤木,起源大陸似乎是亞洲。相反,對八幡草屬族以及整個虎耳草科進行了廣泛的系統發育分析。(Soltis等人,1993,1995,1996),表明八幡草屬的起源中心是北美西部。然而,對於許多被檢查的屬來說,結果是模棱兩可的。例如,唐松毛茛屬的姐妹群是由毛茛屬和鼠尾毛茛屬組成的分支,這兩個屬廣泛分布於北半球和南半球,但人們對其物種之間的系統發育關系知之甚少。同樣,大苞山茱萸的姐妹群也呈環北分布。因此,原點中心是不明確的。對於黃水枝屬來說,在一些系統發育分析中尚未發現有充分支持的姐妹群。因此,還無法確定起源中心。
化石證據將允許進一步評估根據分子數據的系統發育分析推斷的起源中心的生物地理學假說。如上所述,除了山茱萸之外,本文討論的七個分類單元都沒有足夠的化石記錄。
結論
被子植物六屬,八幡草屬、黃水枝屬、唐松毛茛屬、山茱萸大苞山茱萸、楤木屬。楤木屬、夏蠟梅屬以及蕨類植物鐵線蕨(掌葉鐵線蕨)表現出相似的生物地理關系模式:北美東部和西部的物種/族群是姐妹群;這個分支又是東亞物種/族群的姐妹。根據歷史生物地理學的原則(即系統發育拓撲和譜系地理分布之間的一致性表明共享的生物地理歷史),這裏提供的系統發育數據支持了替代假設,即這種眾所周知的植物區系間斷代表了或多或少的分裂第三紀存在連續的中生混交森林群落。然而,我們警告說, 系統發育和地理分布之間的一致性並不一定表明相同的植物地理歷史。涉及東亞、北美東部和北美西部的植物區系間斷可能起源於不同屬的非常不同的地質時期 (例如,Li,1972;Tiffney,1985b),這種現象被稱為「偽一致性」(Cunningham和Collins,1994)。我們最初嘗試使用分子鐘來評估發散時間來評估這種可能性,但並沒有反駁偽同余的假設。 應檢查顯示這種間斷的其他類群的系統發育關系和間斷物種之間的分化時間,並應做出額外的努力來評估假一致性的可能性。
期刊:Molecular Phylogenetics and Evolution
文章標題: The Eastern Asian and Eastern and Western North American Floristic Disjunction: Congruent Phylogenetic Patterns in Seven Diverse Genera
作者資訊:Qiu-Yun Xiang , Douglas E. Soltis, Pamela S. Soltis
原文連結:https://doi.org/10.1006/mpev.1998.0524
文內圖片及封面圖片來源原文











