癌症新型靶点:WRN
WRN是一种具有多种酶活性的多功能的DNA解螺旋酶。WRN表现有DNA依赖的ATP酶活性,ATP依赖的3’→5’DNA解螺旋酶活性,单链DNA退火活性,核酸外切酶活性。
01
Rec Q解螺旋酶概述
解螺旋酶是自然界中一类打开核酸链之间氢键的酶,在生物体中发挥多种重要的功能。RecQ解螺旋酶是自然界的第二大类解螺旋酶,广泛分布在从细菌到人类等众多生物之中,其在生物体DNA代谢、维持基因组稳定过程中发挥着重要的作用。Werner综合征蛋白是RecQ解螺旋酶的一种。在人体中有5种RecQ解螺旋酶,分别是RecQ1、BLM(RecQ2)、WRN(RecQ3)、RecQ4、RecQ5。
RecQ解旋酶属于SF2家族解旋酶,进化过程中从原核到真核生物都是高度保守的,在基因组稳定性的维持中发挥重要功能。
RecQ解旋酶包含三个保守的结构域:如图1所示,RecQ解旋酶独有的Helicase结构域和RecQ羧基末端结构域(RQC),以及解旋酶和核糖核酸DC-末端结构域(HRDC),这个结构域也存在于其他DNA结合蛋白中。除了人类的三个RecQ解旋酶RecQ1、RecQ4和RecQ5,大肠杆菌RecQ三个结构域在其他物种的RecQ同源物中也是保守的。Helicase结构域和RQC结构域形成大肠杆菌RecQ 59kDa的催化核心,在体外该片段可以催化单链DNA(ssDNA)结合并具有3'→5'解旋酶活性,与全长大肠杆菌RecQ的解旋活性基本相同。
其中BLM、WRN、RecQ4功能缺失分别会导致常染色体隐性遗传病布鲁姆综合征(Bloom syndrome,BS),沃纳综合症综合征(Werner syndrome,WS)以及罗斯蒙德-汤姆森综合征(Rothmund-Thomson syndrome,RTS)。这个核心片段的晶体具有四个不同子域的单体:Helicase结构域由两个N末端RecA样亚结构域组成,其中ATP和ssDNA结合位点夹在它们之间,并发挥解旋酶的「马达」功能。解旋酶活性对于体内RecQ解旋酶功能至关重要,并且该结构域中的点突变导致酿酒酵母,小鼠和人类的解旋酶活性钝化。对RQC作用巨大的C端结构域催化核心由Zn2+结合亚结构域和β-发夹螺旋-螺旋(WH)亚结构域组成,它们一起构成dsDNA结合结构域。
在大肠杆菌RecQ中取代Zn2+配位的半胱氨酸残基揭示了Zn2+结合域对稳定DNA结合和蛋白质稳定性是必需的。在BLM和RecQ5中Zn2+配位半胱氨酸的突变产生类似的结果,这表明该功能在进化上是保守的。
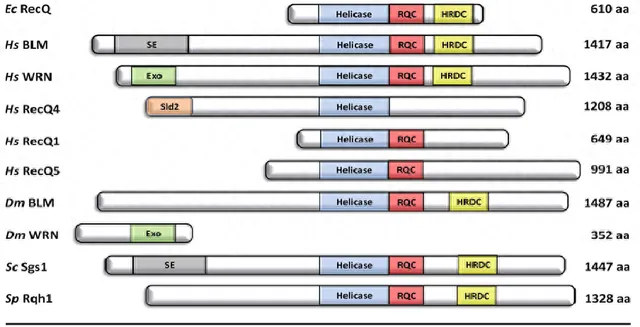
图1 Rec Q解旋酶家族的结构特点
02
Werner综合征
Werner综合征由德国的Werner博士1904年首先报道,又名成人早老症。是一种由于染色体8p11-12发生退行性突变而导致的常染色体隐性遗传性疾病,多见于近亲结婚的子代,以堂兄妹间结婚者的子代居多。
临床表现为包括早衰和各种与早衰相关的继发性症状,包括双眼白内障,皮肤萎缩,身材矮小,头发灰白稀疏。此外患者常会发生II型糖尿病,骨质疏松,动脉粥样硬化和肿瘤。WS患者平均死亡年龄在46-54周岁,其中最主要的死因是恶性肿瘤和心肌梗死。
Werner综合征无治愈方法,主要以对症治疗为主。手术治疗白内障,预防皮肤溃疡,使用降糖药物治疗糖尿病,补充性激素等。
03
WRN基因序列
WRN基因定位在8号染色体8p12,WRN基因基因序列如下:
MSEKKLETTAQQRKCPEWMNVQNKRCAVEERKACVRKSVFEDDLPFLEFTGSIVYSYDASDCSFLSEDISMSLSDGDVVGFDMEWPPLYNRGKLGKVALIQLCVSESKCYLFHVSSMSVFPQGLKMLLENKAVKKAGVGIEGDQWKLLRDFDIKLKNFVELTDVANKKLKCTETWSLNSLVKHLLGKQLLKDKSIRCSNWSKFPLTEDQKLYAATDAYAGFIIYRNLEILDDTVQRFAINKEEEILLSDMNKQLTSISEEVMDLAKHLPHAFSKLENPRRVSILLKDISENLYSLRRMIIGSTNIETELRPSNNLNLLSFEDSTTGGVQQKQIREHEVLIHVEDETWDPTLDHLAKHDGEDVLGNKVERKEDGFEDGVEDNKLKENMERACLMSLDITEHELQILEQQSQEEYLSDIAYKSTEHLSPNDNENDTSYVIESDEDLEMEMLKHLSPNDNENDTSYVIESDEDLEMEMLKSLENLNSGTVEPTHSKCLKMERNLGLPTKEEEEDDENEANEGEEDDDKDFLWPAPNEEQVTCLKMYFGHSSFKPVQWKVIHSVLEERRDNVAVMATGYGKSLCFQYPPVYVGKIGLVISPLISLMEDQVLQLKMSNIPACFLGSAQSENVLTDIKLGKYRIVYVTPEYCSGNMGLLQQLEADIGITLIAVDEAHCISEWGHDFRDSFRKLGSLKTALPMVPIVALTATASSSIREDIVRCLNLRNPQITCTGFDRPNLYLEVRRKTGNILQDLQPFLVKTSSHWEFEGPTIIYCPSRKMTQQVTGELRKLNLSCGTYHAGMSFSTRKDIHHRFVRDEIQCVIATIAFGMGINKADIRQVIHYGAPKDMESYYQEIGRAGRDGLQSSCHVLWAPADINLNRHLLTEIRNEKFRLYKLKMMAKMEKYLHSSRCRRQIILSHFEDKQVQKASLGIMGTEKCCDNCRSRLDHCYSMDDSEDTSWDFGPQAFKLLSAVDILGEKFGIGLPILFLRGSNSQRLADQYRRHSLFGTGKDQTESWWKAFSRQLITEGFLVEVSRYNKFMKICALTKKGRNWLHKANTESQSLILQANEELCPKKLLLPSSKTVSSGTKEHCYNQVPVELSTEKKSNLEKLYSYKPCDKISSGSNISKKSIMVQSPEKAYSSSQPVISAQEQETQIVLYGKLVEARQKHANKMDVPPAILATNKILVDMAKMRPTTVENVKRIDGVSEGKAAMLAPLLEVIKHFCQTNSVQTDLFSSTKPQEEQKTSLVAKNKICTLSQSMAITYSLFQEKKMPLKSIAESRILPLMTIGMHLSQAVKAGCPLDLERAGLTPEVQKIIADVIRNPPVNSDMSKISLIRMLVPENIDTYLIHMAIEILKHGPDSGLQPSCDVNKRRCFPGSEEICSSSKRSKEEVGINTETSSAERKRRLPVWFAKGSDTSKKLMDKTKRGGLFS
04
WRN基因结构
WRN基因跨度超过250 kb,35个外显子,其中34个为编码外显子。编码由1432个氨基酸残基组成、分子质量为160 kD的WRN蛋白(WRNp)。从N端到C端主要分成5部分:核酸外切酶结构域、解螺旋酶(或ATP酶)结构域、RecQ碳末端(RecQ C-terminal,RQC)结构域、解螺旋酶核糖核酸酶D碳末端(helicase-and-ribonucleaseD/Cterminal,HRDC)结构域、核定位信号(nuclear localization signal,NLS)。其中,ATPase结构域和RQC结构域是WRN发挥解旋酶功能的核心。
另外,WRN是其家族成员中唯一拥有核酸外切酶结构域的RecQ蛋白,具有核酸外切酶活性。WRN解旋酶和核酸外切酶结构域既独立又相互协同,以分解多种DNA底物,例如双链DNA、D环、复制叉和气泡结构、G-四链体、Holliday连接、DNA flaps等。

图2 人WRN结构图
05
WRN生物学功能
WRN是一种既可以和DNA结合又可以和其他蛋白质结合的多功能的DNA解螺旋酶,在生物体中发挥多种生物学功能。WRN在DNA复制、DNA重组、DNA损伤修复、基因转录、维持端粒稳定、维持异染色质稳定等生物学过程中发挥重要的作用。
常见的DNA损伤有碱基受到氧化、脱氨基、甲基化等化学修饰而发生改变导致的碱基替换、错配、跨链连接和DNA断裂等。WRN与polδ、polβ、APE1、PARP1等多个参与BER的蛋白在功能上具有相互作用。WRN解旋酶活性能够刺激polβ介导的DNA链置换合成,WRN核酸外切酶和解旋酶活性则可以与polβ协同作用处理3错配。
在DNA双链断裂(double-strand break,DSB)修复中,WRN通过经典的非同源末端连(canonic
non-homologous end-joining,c-NHEJ)修复和HRR发挥作用。c-NHEJ介导的DSB修复在细胞周期中占主导地位。KU70/80异二聚体和DNA-PK形成稳定的复合物,并启动DSB修复;KU70/80异二聚体的直接相互作用可刺激WRN的核酸外切酶活性,有助于处理DSB末端。
当KU70/80缺陷时,DSB修复从c-NHEJ切换为替代型NHEJ(alternative NHEJ,alt-NHEJ)。在alt-NHEJ修复中,DSB末端被PARP和MRE11识别,随后MRE11/RAD50/NBS1(MRN)复合物和CtIP通过切除处理断裂末端。值得注意的是,当WRN被招募到DSB时,它会抑制MRE11和CtIP的招募,并保护断裂的DNA免受5端切除,抑制alt-NHEJ。
HRR由MRN复合物通过MRE11的核酸内切酶和3–5核酸外切酶活性与CtIP启动。细胞周期蛋白依赖性激酶1可磷酸化WRN的丝氨酸1133位点,调节DSB处的MRE11募集来切换HRR和NHEJ。WRN可与DNA复制解旋酶/核酸酶2发生相互作用,促进RPA依赖的DNA末端加工和切除。另外,BRCA1还直接与WRN相互作用并刺激WRN的解旋酶和核酸外切酶活性,促进DNA末端切除。
R环(R-loop)是转录相关基因组不稳定性的主要来源。R环会破坏S期的复制叉进程,导致DNA复制叉的破坏、发生DSB或进入有丝分裂前的不完整复制。研究表明,WRN在限制R环相关基因组不稳定性中发挥了重要作用。ATR激酶是参与识别和稳定停滞复制叉的主要激酶WRN和ATR通路之间存在相互作用;在复制压力下,ATR依赖的WRN磷酸化,充当CHK1激活的介质,促进复制的恢复和防止在停滞的分叉处形成DSB。同时,WRN解旋酶活性对于保护DNA脆弱位点免于断裂至关重要。

图3 WRN蛋白结构域和生物学功能
a−WRN解旋酶的蛋白结构域;b−WRN解旋酶在碱基切除修复长补丁修复途径中的调节作用,WRN识别polβ-DNA复合物并刺激polβ-DNA链置换合成;c−WRN解旋酶在双链断裂修复途径—NHEJ修复中的作用,c-NHEJ是主要的双链断裂修复通路,用粗箭头表示,WRN通过抑制MRE11和CtIP 的募集来促进c-NHEJ并抑制alt-NHEJ;d−WRN解旋酶在双链断裂修复途径—HRR中的作用,WRN与 DNA2、BRCA1发生相互作用,促进DNA末端加工和切除;e−WRN解旋酶在限制R环相关基因组不稳定性中的作用,ATR依赖的WRN磷酸化,可充当CHK1激活的介质,确保R环及时解决,从而确保基因组稳定性。
06
WRN作为癌症靶点
微卫星不稳定性(microsatellite instability,MSI)是一种肿瘤细胞中由错配修复受损或缺陷导致而出现新的微卫星等位基因的现象。MSI可以导致肿瘤细胞基因组进一步紊乱和突变,从而促进恶性肿瘤的发生发展,是公认的重要致癌途径之一。
DNA MMR缺陷的MSI肿瘤细胞在敲除WRN基因或耗尽WRN蛋白后,会发生合成致死效应,导致细胞死亡。MMR缺陷的合成致死相互作用与其他RECQL解旋酶无关,仅与WRN解旋酶活性相关。当WRN解旋酶活性丧失时,MSI的合成致死作用才被激活。
随着WRN可能是MSI恶性肿瘤的合成致死新靶点的发现,基于靶向MSI恶性肿瘤的WRN抑制剂的研究也引起了科研人员和药物研发人员的兴趣。

表1 WRN抑制剂情况
目前处于临床阶段的是Novartis Pharmaceuticals Corp的小分子化药,HRO761, 用于治疗微卫星高度不稳定性结直肠癌,目前处于临床1期。
END
内容源于网络,如有侵权,请联系删除。











