編譯:微科盟 梅子,編輯:微科盟 景行、江舜堯。
微科盟原創微文,歡迎轉發轉載,轉載請註明來源於【轉錄組】公眾號。
導讀
土壤鎘(Cd)汙染日益嚴重,需要有效的修復策略。 本研究調查了外源茉莉酸(JA)對Cd脅迫下東南景天( Sedum alfredii Hance)的影響,旨在提高其植物提取修復效率。研究者首先評估了在Cd濃度為100、300和500μmol/L的環境中添加不同濃度JA的影響 。結果表明, 濃度為1μmol/L的JA最合適。這一濃度透過提高抗氧化酶的活性,有效地降低了ROS產物的水平 。此外,JA還能促進Cd的吸收和積累,同時明顯改善植物的生物量和光合作用。然後, 研究者在300μmol/L Cd脅迫下使用1μmol/L JA處理東南景天,轉錄組分析揭示了一系列JA介導的在響應Cd脅迫中起重要作用的差異表達基因(DEGs)。這些差異表達基因不僅包括JA生物合成和訊號傳導途徑,還包括編碼影響抗氧化系統和光合作用功能的基因,以及與細胞壁合成、金屬螯合和轉運有關的基因 。 本項研究表明,JA處理能顯著增強東南景天對Cd的耐受性和積累能力 ,為植物修復提供了一種前景廣闊的策略,並加深了研究者對植物應對重金屬脅迫的理解。
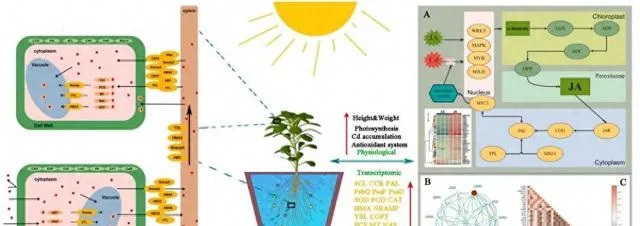
論文ID
原名: Jasmonic acid’s impact on Sedum alfredii growth and cadmium tolerance: A physiological and transcriptomic study
譯名: 透過生理學和轉錄組學分析研究茉莉酸對東南景天生長和耐Cd能力的影響
期刊: Science of the Total Environment
IF: 9.8
發表時間: 2024年1月
通訊作者: 楊文浩
通訊作者單位: 福建農林大學資源與環境學院
DOI號 : 10.1016/j.scitotenv.2024.169939
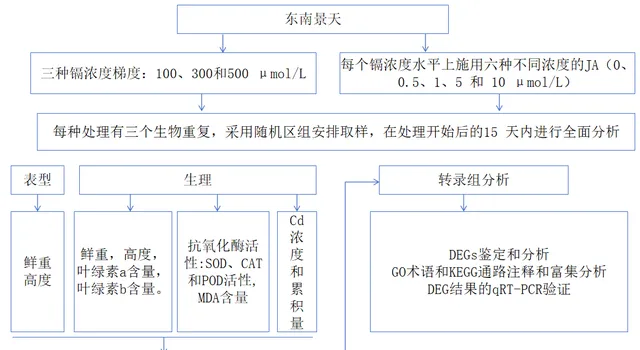
實驗設計
結果
1 施用Cd和JA的植物生長情況
透過植物鮮重和高度的變化,可以清楚地看出JA對東南景天生長的影響。如圖1(A和B)所示, 隨著外源JA濃度的增加,東南景天的鮮重和高度都呈明顯的上升趨勢 。具體來說, 在施用10 umol/L JA的情況下,與施用0μmol/L JA的植株相比,不同Cd濃度下的東南景天鮮重增加了10.12%到29.44%。植株高度也有所增長,增幅從15.67%到7.13%不等 。值得註意的是, 東南景天在1μmol/L JA的條件下達到了生長的頂峰,鮮重達到2.66g,株高達到17.24 cm。
圖1(C和D)顯示了各種處理對東南景天光合色素的影響 。在所有探究的濃度中,Cd的影響主要是抑制作用,隨著Cd濃度的增加,抑制作用逐漸減弱。 與0μmol/L JA處理相比,隨著JA濃度的增加,不同Cd濃度下東南景天的葉綠素水平均呈現先升高後降低的模式 。 在1μmol/L JA濃度下,光合作用和色素沈著的增強效果最好, 尤其是在300μmol/L Cd濃度下,葉綠素a和葉綠素b的濃度分別達到0.27 mg/g和0.11 mg/g。
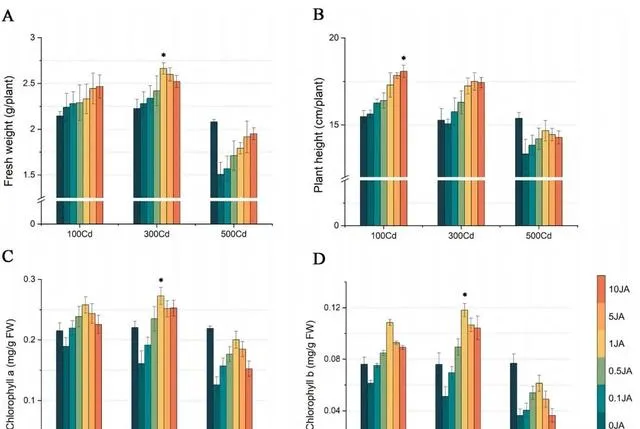
圖1.Cd和JA處理對東南景天生物量和色素沈著的影響。 (A)東南景天的鮮重,(B)東南景天的高度,(C)東南景天的葉綠素a含量,(D)東南景天的葉綠素b含量。
2 施用JA對Cd脅迫下抗氧化酶活性和MDA含量的影響
研究者透過測量東南景天的MDA含量,仔細研究了重金屬引起的氧化應激 (圖2)。與CK相比,在不同的Cd劑量下,MDA含量上升了32.42%至65.16%。然而,加入較低濃度的JA(0.1、0.5和1 μmol/L)後,MDA含量大大減少。更確切地說,在不同的Cd濃度下,1μmol/L JA的用量比0μmol/L JA條件下分別減少了15.38%、16.38%和24.20%。相反,JA劑量越高(5μmol/L和10μmol/L),MDA含量越高,在Cd濃度為500μmol/L時,MDA含量明顯增加,與1μmol/L JA相比,5μmol/L JA和10μmol/L JA處理的MDA含量分別增加了4.38%和12.21%。
同時, 研究者還分析了不同JA處理對東南景天中SOD、CAT和POD活性的影響,目的是減輕Cd誘導的脅迫 (圖2)。結果證明,在所有測試方案中, 較低劑量的JA都能顯著提高抗氧化酶的活性,在1μmol/L JA的情況下達到頂峰 。與不含JA的對應物相比,研究者註意到在各種Cd濃度下,SOD、CAT和POD活性分別提高了27.44%、25.48%和23.46%,38.82%、36.89%和30.61%,以及20.28%、22.90%和20.14%。然而,在5μmol/L JA和10μmol/L JA區間,抗氧化防禦能力逐漸減弱,導致響應收到抑制。
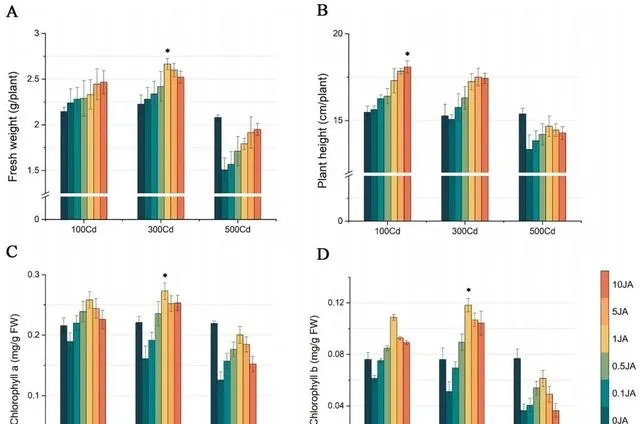
圖2.Cd和JA處理下東南景天的MDA和抗氧化酶活性。
3 Cd脅迫下JA處理對Cd積累的影響
圖3顯示了東南景天顯著的Cd積累能力,0μmol/L JA組的Cd含量分別為4803.7、7508.28和5447.92 mg/kg。引入JA能明顯提高東南景天的Cd積累潛力。 研究者對暴露於100和500μM Cd條件下的東南景天進行實驗,所獲得的數據表明,隨著JA含量的增加,東南景天中的Cd濃度也在持續增加。 在10μmol/L JA條件下,Cd濃度的峰值分別為6640.98mg/kg和5927.60mg/kg,在無JA條件下分別增加了27.68%和8%。在300μM Cd設定中,隨著JA的增加,東南景天中的Cd含量開始上升,但隨後出現下降, 在1μmol/L JA時達到頂峰,濃度為8639.34mg/kg (與0μmol/L JA相比上升了13%)。同時, 研究者對100μM和500μM Cd環境的東南景天研究表明,隨著外源JA濃度的增加,東南景天的Cd積累量也在不斷增加 。在10μmol/L JA條件下,100μM Cd和500μM Cd處理下的東南景天積累值分別為656.41μg/plant和290.42μg/plant,與0μmol/L JA相比,分別大幅增長了1.60倍和2.28倍。 300μM Cd+1μmol/L JA條件是東南景天對Cd積累最有效的條件,達到1009.72μg/plant的峰值(與0μmol/L JA相比提高了1.43倍) 。

圖3.Cd和JA條件下東南景天的Cd濃度和累積量。
4 Cd脅迫下用JA處理的東南景天的轉錄組分析
為了進一步探討Cd和JA處理下東南景天的生理和分子機制,研究者根據東南景天的總體Cd含量及其生理參數,選擇300μM Cd+1μmol/L JA處理組進行轉錄組學分析。 轉錄質控數據見補充材料圖S1和表S2,資料庫註釋數據見圖S2。 研究者透過分析在JA和CK處理的地上部份中發現了7254個DEGs (圖4)。其中, 3503個基因在地上部份上調,3751個基因下調 。相比之下, 根部的DEGs數量更為顯著,共有10399個。其中絕大多數,特別是9554個基因被上調,只有845個基因被下調 。所有DEG都符合絕對log2(fold change)≥1和 P 值<0.05的標準。
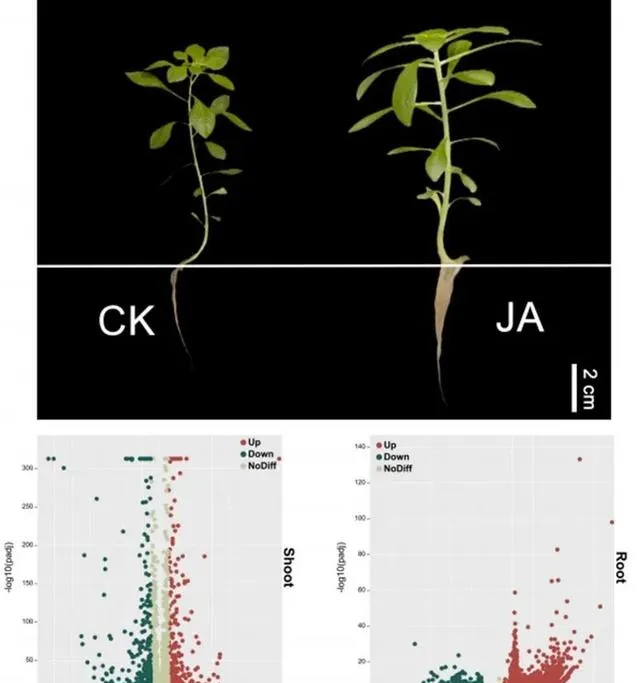
圖4.東南景天在Cd和JA條件下的生長和差異基因的表達。
5 GO術語和KEGG通路註釋和富集分析
在東南景天對Cd和JA處理的響應中,GO註釋分析(圖S3)揭示了DEGs在三個本體中的分布:生物過程(BP)、分子功能(MF)和細胞組分(CC)。 從TOP30 GO術語中,研究者可以觀察到以下趨勢:在用JA處理的東南景天的地上部份中,有23個GO術語被歸入CC類別 , 主要與"細胞膜和細胞壁"基因有關。在BP和CC兩類中,最富集的GO術語與"光合作用"和"金屬和陽離子轉運"有關 。而 在根分析中,在TOP30的GO術語中,有14個術語被歸入生物過程(BP)類別,主要與氮化合物和胺的生物合成有關。13個術語被歸入CC類,主要集中在與核糖體有關的結構上。CC類別還包括結構功能。
為了深入研究JA促進Cd脅迫下東南景天中DEGs的生物學特征,研究者利用KEGG通路資料庫進行了通路富集分析 (圖S1)。 結果表明,東南景天的KEGG代謝通路包括細胞組織結構和分子運輸、訊息傳遞、胺基酸代謝、脂肪酸及其衍生物代謝和光合作用等多個方面 。具體來說, 在地上部份,最主要的代謝途徑是過氧物酶體、內吞作用、碳代謝和植物與病原體的交互作用。相比之下,根部的主要代謝途徑包括裂殖體、RNA運輸、碳代謝和脂肪酸代 謝。
6 DEG結果的qRT-PCR驗證
為了證實轉錄組測序的準確性,研究者對來自芽和根的四個關鍵基因進行了qPCR檢測,這些基因與多種Cd響應途徑密不可分 (圖S4)。所選基因主要與細胞壁合成、金屬螯合和運輸過程有關。它們的表達譜與Illumina測序結果非常吻合,進一步驗證了DEG分析的可靠性。
討論
1 JA在Cd脅迫下促進東南景天的生長
在本研究中,研究者觀察到JA對東南景天在Cd脅迫下的光合作用有積極影響,表現為葉綠素-a和葉綠素-b的含量顯著增加 (圖1)。這與芽中的許多DEGs有關,包括參與光合系統I和II、光合電子傳遞鏈、F型ATP合酶和光合葉綠素-蛋白質復合體的DEGs(圖5)。在光合作用過程中,光系統I和II被證明共同驅動電子傳遞鏈,協同完成光合電子傳遞。它們將光能轉化為生長所需的能量和有機化合物,支持植物的生長和發育。然而,Cd脅迫會阻礙這一過程,降低光系統I的活性,增加ROS的產生,從而影響卡爾文迴圈和其他生物合成途徑。 本研究發現,JA可透過調節PSII和PSI的基因表現,增強光合系統的穩定性和活性,這有助於植物更好地適應外部環境脅迫,保持健康的生長和發育 。 研究者還觀察到,JA影響F型ATP酶多個亞基的表達 。這種酶是一種多亞基蛋白質復合物,透過質子動力催化ATP的合成或水解。 本研究表明,外源JA可以影響這種酶的各種亞基的表達,維持細胞內的pH平衡和能量供應,從而確保細胞的完整性 。這有助於植物在面臨Cd脅迫時維持正常的細胞功能,從而促進其生長和適應力。

圖5.Cd和JA條件下與東南景天光合作用相關的DEGs分析。
2 JA提高了東南景天對Cd脅迫的抗性
2.1 JA透過細胞壁提供額外的保護,降低Cd的毒性
Cd2+透過各種途徑進入植物體內,對植物的生長和發育產生不利影響(圖6)。植物細胞壁是最初的屏障,具有固定和捕獲Cd2+的能力,從而減少其在細胞內的積累。 在植物的地上部份,研究者觀察到參與木質素生物合成的CCR(DN81890_c3_g5和DN78139_c1_g1)表達上調,這可能會增強細胞壁強度並限制Cd2+的滲透 。此外,其他一些與木質素合成相關的實體,包括PAL(DN81699_c1_g5)和4CL(DN86000_c4_g4)的表達上升,也被證明有助於增加細胞壁密度,阻礙Cd2+的滲透。FAR(DN83112_c3_g1)和BGLH(DN80382_c1_g4)表達水平的升高可能會影響細胞壁的結構和穩定性。與地上部份相比, 根主要涉及與多糖和果膠合成有關的關鍵因子 ,如Cals(DN87352_c0_g1)和XTH(DN84935_c0_g1)。這些實體透過最佳化細胞壁的多糖結構,增強了對Cd的抗性。此外,PE可能透過調節細胞壁中果膠的合成來調節其穩定性和阻斷Cd2+的能力。JA似乎可以透過上調CHI(DN83308_c1_g1)和EXP(DN145391_c0_g1)來增加細胞壁的可塑性和擴張性,以對抗Cd脅迫。 研究者還註意到,EXT(DN75005_c0_g2)家族的下調有可能導致細胞壁更具彈性和適應力,更適於承受外部壓力,而不僅僅是增加硬度以抵禦Cd的侵入 。同時,LAC(DN84135_c1_g2)和CAD(DN141324_c0_g1)參與了木質素合成的調控。CAD表達的增加意味著植物在應對Cd脅迫時強化細胞壁結構的策略,而LAC表達的減少則表明特定木質素生物合成途徑可能受到限制。預計這種平衡可保持細胞壁的強度和柔韌性,而不會過度硬化。
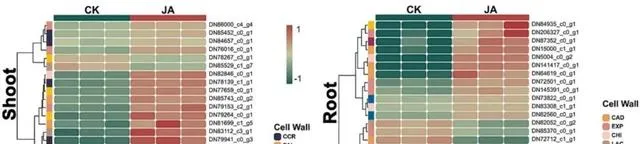
圖6.在Cd和JA條件下參與細胞壁合成的DEGs。
總之, 研究者認為外源JA顯著上調了一系列與細胞壁合成和修飾相關的實體,增強了植物對Cd脅迫的抵抗力,說明JA在細胞壁響應機制的調控中發揮著關鍵作用。 這些修飾將導致細胞壁更致密、更堅固,從而有效抑制Cd2+的滲透。
2.2 外源JA可透過抗氧化系統提高東南景天的抗逆性
植物前進演化出了非酶和酶的防禦機制來抵禦氧化應激,以應對具有挑戰性的環境。先前的研究表明,植物激素JA能夠刺激SOD、CAT和APX等抗氧化酶的合成,同時抑制ROS的產生,從而增強植物對Cd的耐受性。最新研究發現,在Cd脅迫下,JA合成基因AtAOS可刺激內源JA的產生,從而進一步抑制Cd引發的ROS生成,減輕Cd對植物生長的抑制作用,增強植物對Cd的抗性,本研究證實了這些發現(圖7)。 研究者發現,添加外源JA能顯著提高東南景天中參與抗氧化系統的幾個關鍵基因(包括SOD、APX、GPX、PRX、CAT和GR)的表達水平,這說明JA在調節植物對Cd脅迫的響應,尤其是在建立和增強抗氧化防禦系統方面發揮著核心作用 。總之,JA可透過多種途徑增強東南景天的抗氧化防禦機制,包括上調參與清除ROS的基因和促進建立更復雜的抗氧化網路。這有助於植物在Cd脅迫下保持穩定和有生命力的內部環境,提高其抗性。
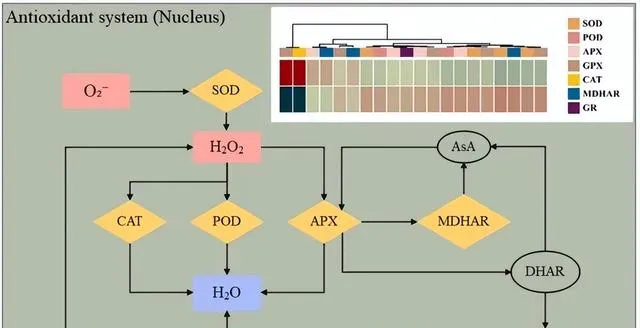
圖7.在Cd和JA條件下參與抗氧化系統的DEGs。 O2-(超氧陰離子)、pO2(過氧化氫)、pO(水)、SOD(超氧化物歧化酶)、CAT(過氧化氫酶)、POD(過氧化物酶)、APX(抗壞血酸過氧化物酶)、GPX(谷胱甘肽過氧化物酶)、ASA(抗壞血酸)、MDHAR(單去氫抗壞血酸還原酶)、DHAR(去氫抗壞血酸還原酶)、GSH(谷胱甘肽)、GR(谷胱甘肽還原酶)。
3 JA透過調節金屬螯合和轉運蛋白促進東南景天對Cd的吸收和積累
通常情況下,當Cd2+穿過細胞壁進入細胞時,植物會啟動一系列生化反應來降低Cd的毒性。最主要的反應是螯合,即利用小分子配體與Cd2+形成穩定的復合物。 在本研究中,研究者觀察到外源JA顯著調節了幾個編碼與金屬螯合相關的蛋白質的基因的表達(圖8),揭示了JA增強了東南景天螯合Cd2+的能力 。 研究者首先註意到PCs(DN77959_c1_g1)和MT(DN76429_c1_g3)表達的上調 。由PCS基因編碼的蛋白植物螯合素能促進植物螯合素與Cd2+復合物的形成,有效降低細胞中遊離Cd2+的濃度,從而減輕Cd的毒性。研究表明,當過表達來自苧麻( Boehmeria nivea )的新型植物螯合酶基因BnPCS1時,阿拉伯芥對Cd2+的耐受、積累和轉運能力都會增強。金屬硫蛋白(MT)編碼金屬硫蛋白,有助於維持細胞中金屬離子的平衡和解毒。雖然SaMT3主要與鋅和銅形成復合物,但Zhao等人的最新研究發現,東南景天中的SaMT3具有Cd螯合能力。透過異源表達煙草金屬硫蛋白基因NtMT2F,阿拉伯芥對Cd脅迫的耐受性明顯增強。此外, 研究者還觀察到NAS(DN85360_c2_g5)、GST(DN82774_c0_g3)、Cys(DN148504_c0_g1)和OAS(DN218485_c0_g1)的協調上調。 值得註意的是, NAS透過煙堿胺的合成發揮了重要的金屬螯合劑作用。它的上調表明,JA增強了金屬螯合過程,從而減輕了地上部份的Cd毒性,並限制了根系對Cd2+的吸收 。另一方面,Cys和OAS是編碼參與半胱胺酸合成(GSH的前體)功能的關鍵基因。因此,這些基因的上調很可能是放大GSH合成途徑的關鍵,從而有效地管理Cd的封存和儲存。
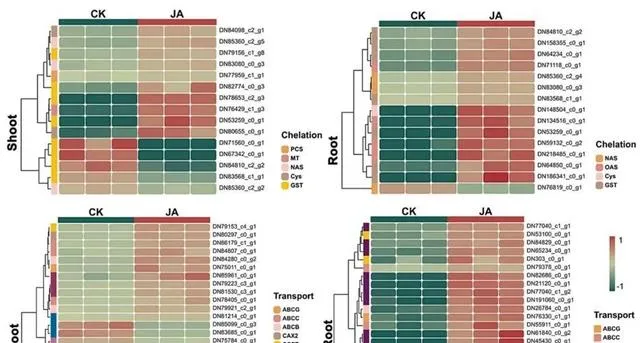
圖8.Cd和JA條件下參與金屬螯合和運輸的DEGs。
金屬轉運蛋白在高積累植物吸收和轉運金屬的過程中起著至關重要的作用。盡管Cd不是植物的必需元素,而且在植物系統中也沒有建立專門的轉運途徑,但轉錄組數據顯示,外源JA能夠透過增強其他二價離子通道(如Zn、Ca或Fe通道)中Cd2+的轉運能力來調節東南景天的Cd積累。 為了系統分析這些基因在Cd管理中的作用,研究者將它們分為三大功能類別:(1)液泡中的Cd儲存;(2)木質部和細胞質之間的Cd相互運輸;(3)從根到芽的Cd長距離運輸。
液泡是植物細胞中Cd2+的主要儲存場所,其中VTL在Cd2+從細胞質轉移到液泡的過程中發揮著關鍵作用。VTL是一種關鍵的鐵轉運蛋白,控制著Fe2+在細胞質和液泡之間的轉運。有趣的是,雖然OsVTL1和OsVTL2已被證明參與了Fe2+的運輸,但它們的過表達導致Cd脅迫下Cd在液泡中大量積累,這表明它們可能有助於螯合Cd和維持植物體內的金屬平衡。 本研究結果表明,JA可選擇性地調節VTL家族基因的表達,特別是透過上調VTL1(DN68490_c0_g1)在地上部份的表達,而降低VTL1(DN68490_c0_g1和DN81816_c1_g1)在根部的表達 。這種差異表達可能揭示了JA在調節Cd反應中的多層作用。例如,地上部份VTL的上調可能會促進更多的Cd被螯合到液泡中,從而增強植物對Cd脅迫的耐受性,從而降低Cd的毒性。同時, 根系中VTL1的下調可能是一種負反饋機制,旨在減少Cd的吸收和/或積累,從而保護根系免受Cd脅迫的損害。
在東南景天中,細胞質和木質部之間的Cd雙向運輸在調節Cd在植物各部份的分布中發揮著重要作用。CAX、NRAMP和HMA蛋白家族在這一過程中發揮了核心作用,確保了Cd的高效和平衡流動。其中,CAX是一種鈣離子轉運蛋白,對維持細胞內Ca2+的平衡和調節植物體內Cd2+的轉運有著至關重要的影響。Zhang等人在東南景天中發現的SaCAX2h蛋白已被證明可調節Cd從細胞質到木質部的流動。值得註意的是,CAX的表達在地上部份和根部都受到外源JA的調控,這意味著 JA有助於提高東南景天對Cd的轉運能力 。NRAMP基因家族被廣泛認為能介導Cd2+和Fe2+的轉運。 根據研究者的觀察,NRAMP2和NRAMP5在Cd在細胞質和木質部之間的雙向轉運過程中表現出顯著的表達差異。 具體來說,N RAMP5限制了Cd從細胞質向木質部的移動,而NRAMP2則促進了這一過程 ,這與Tian等人和Ge等人的研究結果一致。雖然HMA蛋白家族最初主要與Cu2+和Zn2+的轉運有關,但也有研究表明它也參與了Cd2+的轉運。特別值得註意的是, 研究者觀察到HMA5基因在地上部份和根部都出現了上調。這可能表明外源JA透過調節HMA5影響了Cd在細胞質和木質部之間的轉運,進一步最佳化了Cd在植物體內的分布 。
除了Cd在液泡和細胞質中的運輸外,Cd透過木質部從根部向地上部份的移動也是一個至關重要的過程,這一過程確保了Cd在植物不同部位的有效分布,而YSL和ABC轉運家族在促進這一過程中發揮了關鍵作用。黃色條紋樣(YSL)轉運體屬於寡肽轉運體(OPT)家族,主要負責在植物細胞膜上轉運金屬-植物苷元。研究表明,YSL家族參與了Cd的吸收及其從根到芽的長距離運輸。 在本研究中,研究者觀察到YSL12(DN82418_c1_g2)基因表現上調,這表明該基因在外源JA的影響下可能更加活躍,從而增強了其在金屬-植物苷元轉運中的作用 。這可能會增加Cd-植物苷元的吸收,進一步影響Cd2+的整體運輸和分布。同時,YSL6(DN75784_c0_g1)表達的下調可能表明其在這一過程中的功能受到了限制。考慮到已知YSL6參與了Cd2+的長距離轉運,其表達的降低可能為這一Cd2+轉運途徑提供了一種特有的保護機制。此外,ABC轉運體蛋白家族也在木質部內轉運Cd2+的過程中發揮作用。在這個家族中,ABCB、ABCC和ABCG亞家族被認為最直接參與Cd-植物苷元的轉運。AtABCC1和AtABCC2在調節Cd從根到芽的轉移過程中起著至關重要的作用。同樣,OsABCG36和OsABCG43的高表達與水稻木質部內的Cd遷移有關。Naaz等人在大豆中發現,ABCB、ABCC和ABCG運輸蛋白能夠調節Cd2+的吸收和分布,幫助維持植物體內的Cd平衡。這與本研究結果不謀而合,因為 研究者檢測到,在外源JA的作用下,東南景天的地上部份和根部ABCB、ABCC和ABCG的表達都出現了上調,這證實了ABC轉運蛋白家族在透過木質部將Cd從根部轉運到嫩枝的過程中發揮了關鍵作用。
此外, 研究者還觀察到東南景天芽中COPT和Nipa的表達發生了變化 。這些是Cu2+和Mg2+的轉運體,大量研究表明,Cd2+透過模仿Cu和Mg離子的化學性質,從而"欺騙"這些轉運體,透過這些通道滲入植物細胞。 在本研究中,JA的引入明顯增加了這些通道的表達量,促進了Cd2+向細胞內的更大遷移。
總之, JA在控制東南景天中的Cd2+轉運方面發揮著明確的核心調節作用 (圖9)。這種作用超出了對一個或少數幾個分子通道的影響, 它協調了跨膜轉運蛋白家族的廣泛範圍,並突出了其在植物應對Cd脅迫過程中的多方面功能 。最初, JA透過引導幾個家族(包括CAX、VTL、COPT、Nipa和HMA)的表達動態,引導Cd2+在細胞內外的移動和儲存 。例如,它透過促進金屬離子向液泡的轉運來支持維持細胞內離子平衡。這種作用既能保持植物細胞結構的完整性,又能減輕Cd過度積累的毒性影響。 隨後,JA透過增強NRAMP和YSL家族選擇性基因的活性,控制植物根部對Cd的吸收以及Cd在根部和葉片之間的長距離轉運,從而在Cd誘導的脅迫下保護植物的生理結構和生長潛力 。 最後,JA還能刺激ABC家族的特定元素,加強植物的Cd排斥機制。 這些基本功能在Cd的吸收和清除中起著關鍵作用,確保了細胞內外物質的無縫運輸,從而保護植物免受Cd脅迫的有害影響。
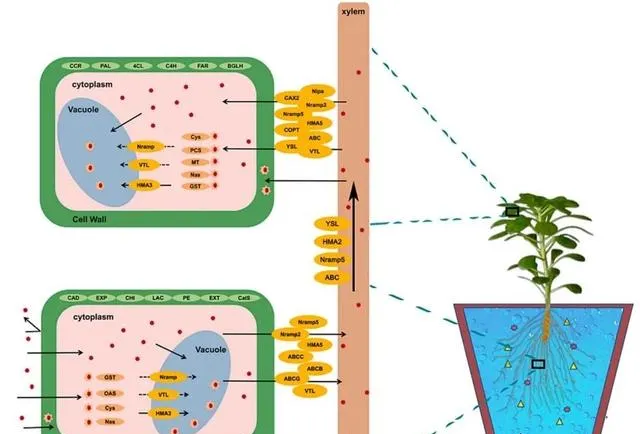
圖9.在Cd和JA條件下參與Cd2+的DEGs示意圖。
東南景天對Cd的吸收和積累受外源JA的顯著影響。在Cd的吸收過程中,當Cd2+進入細胞質時,會與GST、NAS、CYS和PCS等蛋白質形成穩定的螯合物,提供主要的吸收通道。隨後,CAX家族在Cd的細胞內積累過程中發揮了重要作用,促進了Cd從細胞質向液泡的移動 。 同時,NRAMP和HMA家族為Cd在細胞質和木質部之間的迴圈提供了核心途徑,進一步加強了Cd在植物體內的積累 。 在木質部,Cd的向上運輸主要由YSL和ABC家族成員完成 。值得註意的是,在這些關鍵過程中, HMA和NRAMP家族都受到JA的調控,這突出表明了JA在整個Cd吸收和積累機制中的決定性作用。
4 JA生物合成和訊號傳導在東南景天的生長和防禦中的關鍵作用
在Cd脅迫條件下,植物體內JA的合成和訊息傳遞對植物的生長、抗逆性以及Cd的吸收和積累至關重要。當植物面臨Cd脅迫時,它們的最初反應是產生大量活性氧(ROS)。ROS可被視為一種細胞警報,表明細胞內出現了脅迫。為了有效應對這種情況,植物會啟用各種號誌機制。在這一過程中,WRKY、bHLH、MAPK和MYB等轉錄因子起著關鍵作用(圖10A)。特別是在有外源JA的情況下,這些轉錄因子的活性會增強。例如,已知WRKY46可調控JA相關基因對Cd脅迫的反應,MAPK訊號通路被啟用後可進一步調控JA相關基因。 本研究表明,Cd脅迫下轉錄因子WRKY46、MAPK3和MYB的表達上調,而WRKY4、WRKY50和bHLp5的表達下調。這種變化強調了外源JA對特定訊號通路的調控影響,可能會增強東南景天內源JA的生物合成 。
由Cd誘導的轉錄因子在外源JA的存在下表現出對JA生物合成的積極調控作用 (圖10A和B)。具體而言,植物體內LOX、AOS、AOC和OPR3基因的表達量顯著增加,表明外源JA對JA生物合成有促進作用。 JA及其衍生物(如JA-Ile)在生物合成後可作為訊號分子,啟動特定的轉錄因子。這些轉錄因子進一步調節與JA反應相關的基因的表達,從而影響抗逆相關蛋白的合成 。 當JA-Ile與COI1結合時,JAZ蛋白降解,釋放並啟用MYC2轉錄因子。隨後,MYC2轉移到細胞核中,調節與JA反應相關的基因的表達 。 最終,MYC2作為一個關鍵調控因子,協調各種機制以減輕Cd脅迫引起的氧化損傷、提高抗氧化酶的產量、促進東南景天的生長並改善Cd的吸收和富集 (圖10C)。
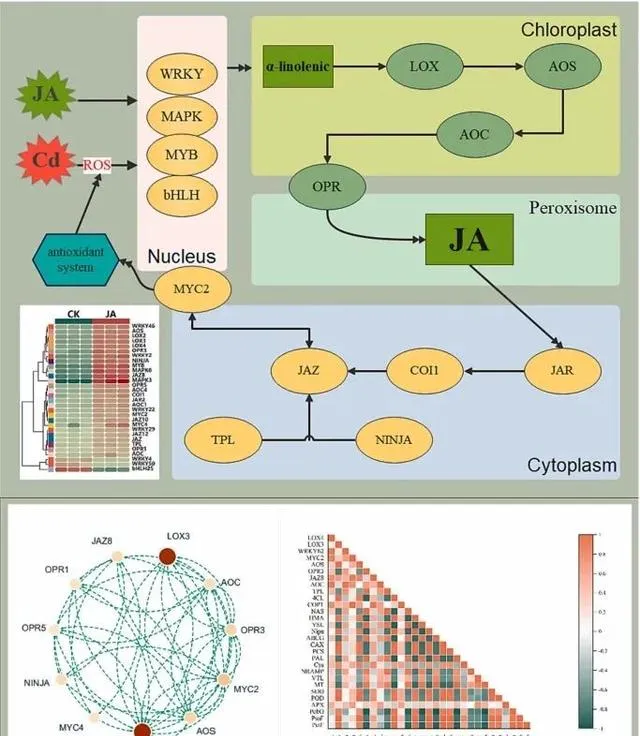
圖10.JA在Cd脅迫中的調控作用。 (A)JA生物合成和訊號傳導機制;(B)JA通路的蛋白質交互作用網路;(C)熱圖顯示了東南景天中與JA、光合作用、抗氧化劑和Cd吸收/積累有關的基因相關性。
結論
本研究透過比較分析外源JA處理對Cd脅迫下東南景天的生理效應和轉錄組效應,揭示了JA在調節植物對Cd脅迫的響應中發揮的多種作用 。 本研究結果表明,施用JA能夠減輕Cd脅迫對植物生長的不利影響,透過最佳化光合作用促進正常的生理條件和生長 。此外, JA在增強植物抗氧化防禦機制方面發揮了重要作用,透過調節編碼抗氧化酶的基因的表達,幫助清除ROS,從而維護細胞內的穩定和秩序 。更重要的是, JA處理明顯減少了Cd在植物體內的積累,表明JA有可能成為減輕Cd脅迫影響的有效策略 。 轉錄組分析發現了一系列在JA介導的Cd脅迫響應中起作用的DEGs, 為了解JA如何透過調控基因表現來操縱植物響應提供了新的視角。 它涵蓋了編碼參與抗氧化系統和光合作用功能的基因,並涉及細胞壁合成、金屬螯合和轉運等多個方面,展示了一個多層次、錯綜復雜的調控網路 。總之,本研究深入揭示了JA在東南景天的Cd脅迫響應中的作用,為其在植物生物技術和環境修復中的套用前景鋪平了道路。











