
Ram C. Poudel 作者
米凯 译者
梁金 审校
我们习惯于从不同的视角看待物质、生命和演化。本文受到经典力学和热力学的启发,提出一个简单但统一的理论框架。该框架将牛顿第三运动定律推广到包含生命和演化的领域。这种广义的作用-反作用 (action-reaction) 关系包含了尺度和时间参数。这种推广有助于解释为什么生命是一个非平衡系统。生命可以超越物质的作用-反作用对称定律。这里将生命定义为一个开放系统,能够自我感知能量状态和环境的时间轨迹。本文是 Philosophy Transanctions of The Royal Socienty 「热力学2.0:连接自然科学和社会科学 (第一部分) 」主题特刊的一部分。
关键词:生命演化,热力学,非平衡物理,作用-反作用
1.引言
物质科学以牛顿定律为基础得以成形;然而,生命和演化的科学仍在发展之中。物质是可观测宇宙的现实基础,以空间、质量和体积为特征。虽然生命和演化的定义有许多种,但我们尚未达成共识。根据美国国家航空航天局 (NASA) 的定义:「生命是一种能够进行自我维持的化学系统,具备达尔文演化论的能力」。自然选择是达尔文提出的一种解释演化的机制。自然选择并不是物理层面的解释。然而,它帮助人类知识从过去的「神选」时代进步到「自然选择」时代。根据斯宾诺莎的观点:Deus sive Natura (神即自然,自然即神) *;神在本质上是与自然等价的。科学——尤其是物理学——要求以物理实体如能量、力及其相互作用来解释。据我们所知,现有文献中对演化的科学解释是不足的。我们提出了一个基于经典力学中场的概念的统一理论框架来研究物质、生命和演化。
*Deus sive Natura(拉丁文,神即自然,自然即神)是与斯宾诺莎哲学思想相关的一个重要概念。斯宾诺莎是17世纪的荷兰哲学家,他提出了一种非常自然主义的关于神、世界、人和知识的观点,这些观点为他的道德哲学、政治思想以及对宗教和道德传统的批判提供了基础。
什么是生命?自从人类从四足动物演化到两足动物以来,我们可能一直在思考这个问题。这是一个尚未解决的问题,也是一个活跃的研究领域。生命科学的发展已经持续了很长时间。东西方哲学对生命有不同的解释。东方教义强调生命是自然的一部分,提倡与自然和谐共处。「Vasudhaiva Kutumbakam」 (字面意思是「世界大同」) 是东方文明中的一个显著的本体论。在西方文化中,生命的主导哲学是「个人主义」,认为「生命是对权力的斗争」。
*Vasudhaiva Kutumbakam 是一个源自古印度的哲学概念,其含义为「世界大同」,这一概念强调了全人类的统一和相互联系。这个短语由梵文词汇「Vasudha」(地球)和「Kutumbakam」(家庭)组成,它传达了一种世界观,即整个世界是一个大家庭,所有生命都是相互连接的。
在缺乏统一科学的情况下,生命对人们来说就像盲人摸象——每个人都根据自己的背景、有限的经验和知识来解释生命是什么。
观察生命有许多不同的视角:生命作为化学;生命作为物理;生命作为信息;生命作为流动组织。我们知道,生命的研究属于生物学范畴。但是,生命知识的的片面理解并不能帮助我们找到科学的统一性。我们需要一个视角,来认识生命在其整体上的多面性和多维性。能量是科学的语言,不受化学、物理和生物学等学科界限的限制。系统状态的这一标量函数可以被用来对物质、生命和演化进行综合性研究。
本文提出一种在能量的连贯框架下理解生命、物质和演化的方法。能量是时空界限的主人,而不是奴隶。然而,在本文中,我们坚持采用场的方法来构建和扩展知识,并遵循经典物理学的方法,以便使本文的内容对一般读者来说易于理解。
一门统一的科学要求将物质和生命在核心层面联系起来。在根本层面上,科学必须是独一无二的。我们不希望物质有一门科学,生命又有另一门科学。在最基本的层面上,生命的任何定律或规则必须与物质的定律或规则相对应,而不是相反。 自然界中普遍存在着标 度无关 的模式 (scale-free pattern) ,这为非生命体与生命体之间的连续性提供了令人信服的证据 。在跨越分界的数据中,普遍存在一种模式,即数据以S形方式累积分布,因此大多数情况下遵循幂律分布。我们可以将这种模式与能量分散过程联系起来。能量及其守恒是热力学中的一个重要概念。能量可能是自然科学和社会科学各学科之间的统一纽带。我们研究热力学中的能量及其转化。本文的主要目的是通过一个更广义的理论框架来扩展热力学的边界,以涵盖生命科学。
2. 牛顿定律与生命科学
恩斯特·马赫*认为牛顿第三定律是经典物理学的基石。正如我们所认为的那样,牛顿三定律是物质的定律,而不是生命的定律。然而,这三定律并不是独立的,它们为理解生命的一些关键特性提供了重要见解。生命可能超越牛顿定律的范畴。在这里,我们从物质的角度出发,总结了我们对生命的关键——即生命的定义——的探索。
*恩斯特·马赫(Ernst Mach)是一位奥地利-捷克物理学家和哲学家,他因其在光学、力学和波动力学方面的工作而闻名。他最著名的贡献是推导出了「马赫原理」,该原理通过假设宇宙中所有的质量都以某种方式相互连接来解释惯性现象。马赫的名字常常通过马赫数被人们所记忆,马赫数与超音速相关联。
我们尝试用牛顿第三定律来阐明这一观点。牛顿第三定律指出,作用力和反作用力是大小相等、方向相反的。对于两个物体A和B之间的因果相互作用,作用力和反作用力 (分别用R表示) 遵循以下规则:
R AB = -R BA (2.1)
作用力R BA 和反作用力R AB 分别作用在两个不同的物体上。第三定律在数学上由方程 (2.1) 表示,它隐含地假设作用力和反作用力在没有时间滞后的情况下同时发生。一些文献给物质定律的这一独特方面起了一个名字: 物自在主义 (object egotism) 。 物体只对瞬间作用于它们的影响做出反应 。但这种物自在主义可能不适用于生命;生命可以超越物自在主义。生命体具有操纵反应以对自己有利的能力。此外,人类——也许任何生命形式——都可以在相同的时间尺度上采取自发 (自主) 行动,或者可能领先或滞后于反应。
当一个定律要求作用-反作用对称时,自然会想知道在哪些情况下作用不等于反作用。确实,我们需要对物质的经典物理学进行一种跳出常规的思考。根据玻尔在数学结构中的对应原理 (correspondence principle) ,我意识到作用-反作用不对称可能与远离平衡态的系统有关,比如生命。如果能将物质的定律推广到生命领域不是很美妙吗?这正是一个统一科学应该努力追求的目标。
*Bohr’s correspondence principle,尼尔斯·玻尔的对应原理是量子力学中的一个基本原则,它指出在某些特定条件下,量子系统的行为应该能够近似地对应到经典物理系统的预测。作者使用这个原理来说明,在数学和物理学的框架内,可能存在一种情况,即作用和反作用的不对称性与非平衡系统(如生命系统)有关。
当我分析一座坐落在柱子上的桥梁时,首次遇到了一个非直观的作用与反作用模式。这座桥梁设计用于尼泊尔等地震多发地区,柱子在承受荷载时可以略微屈服。我对分析结果感到困惑,于是开始思考牛顿第三定律是否具有普遍性,即是否既适用于非生命体 (如物质) 也适用于生命体。自然界中是什么要求作用在两个不同物体上的力和反作用力必须相互平衡?是对称性还是某种守恒定律?这是否是受稳态描述影响的一种思维方式,旨在使物理问题变得简单且易于管理?确实,我们需要先学会走路,然后再努力奔跑。这种基于平衡的解释可能仅对物质有效,而对生命则不然。人们普遍认为生命是一个远离平衡态的过程。对于一个远离平衡态的系统,应该考虑额外的力 (或能量) 来恢复作用-反作用相等的原理。换句话说,像生命这样的非平衡态系统拥有一种在经典物理学中不常见的能量 (或力) 。这听起来可能有些牵强,但这种无法解释的能量可能是生命科学的基石。
本文是从物质的角度探索生命的关键。因此,让我们以生物细胞 (生命的基本构建块) 为参考,来审视物质的三大定律。生命由细胞及其所处的环境组成,是自然界更大生态系统的一部分。
细胞是一个在环境中由边界定义的系统。细胞具有内部能量产生源 (或动力源和某种形式的储存器——如线粒体) ,它可以根据自身和环境的约束随意处置或调节这些能量。细胞产生能量以驱动生命的各种功能和机制。细胞利用能量进行基因组的转录和翻译等功能。活细胞可以自身改变其能量状态,或者对外部力作出反应。因此,我们可以声称牛顿第一定律在细胞层面并不成立。
值得强调的是,我们在这里处理的是一个由细胞边界和环境定义的宏观系统。如果我们深入到更小的尺度,即细胞的组成部分,那么我们主要处理的是物质而不是生命。这也可能强化了一个事实,即牛顿第一定律不足以处理生命及其轨迹。
因此,生命是由边界定义的,并拥有像麦克斯韦妖*开启大门的内在功率。每个生命体都拥有内在的能量来源,或者通过调动宿主或环境产生能量并行使功率 (power) **的能力。边界是生命固有的特征。我们可以定义两种类型的 功率 来源:边界「内部」和「外部」的 功率 来源。「外部」 功率 是指在边界墙外来源的 功率 。在经典物质研究中,我们将力区分为体力 (body forces) 和表面力 (surface forces) ***。重力是一种体力,而压力是表面力的一个例子。社会潮流和时尚可以被视为表面力的类似物。
*麦克斯韦妖(Maxwell’s Demon)是1871年由英国物理学家詹姆斯·麦克斯韦提出的一种思想实验,用来探讨热力学第二定律的可能性和局限性。在这个假想实验中,一个容器被分成两个部分,中间有一个可以由「妖」控制的小门。这个「妖」能够探测并控制单个分子的运动,它可以将速度较快的分子(被认为是「热」分子)放到容器的一边,而将速度较慢的分子(被认为是「冷」分子)放到另一边。这样,容器的一侧温度会升高,另一侧温度会降低,从而可以利用温差做功,这似乎违反了热力学第二定律,因为熵似乎减少了。后续的研究表明,麦克斯韦妖并不违反热力学第二定律。信息的处理,包括信息的擦除,需要消耗能量,这会导致系统的熵增加,从而不违反热力学第二定律。参看【麦克斯韦妖文章】。
***body forces and surface forces:体力(body force)是指作用于物体内部每一个点的力,与物体的表面积无关。表面力(surface force)则作用于物体的表面或边界上,通常与物体的表面积成正比。
生物体能够感知自身的能量状态,并因此渴望随着时间的推移向更高能量状态发展。因此,「功率」更适合描述生命和生物体的动力学 。尼采和罗素早先强调了生命的这一特性。遵循物质研究的传统,我们可以将与生命相关的功率分为两类:(a) 体功率和 (b) 表面 功率 。体 功率 是生物实体的一个定义特征。由于意识到并能够访问这种 功率 ,生物体可以与其他生物体和环境进行自发性和因果性互动。一般来说,物质被认为不具备自发行动的能力;然而,在放射性物质的情况,我们可以观察到这种行动。在根本层面上,可能没有明确的边界来区分物质和生命。
功率 关系和层级制度似乎调节着对能量状态有意识的生物体的行为和反应。 功率 仅仅是我们称之为能量的系统状态函数的总导数。在下一节中,我们将探讨与生命相关的能量和 功率 。
*全导数( total derivative ),系统能量状态函数关于时间或其他相关变量的变化率。
3. 能量和功率
测量是科学的基础。与物质相关的测量大多涉及空间、时间和能量 (或力) 。能量是科学的基本概念之一,大多数测量都是基于能量或其导数,如力和功率。然而,与生命相关的能量仍然是一个谜。
理解人类的能量 (及功率) 以及如何利用它更好地服务人类和自然,应该是21世纪科学和工程的主要目标之一。我们在理解支持生命的物质上花费了太多时间和资源。最近,人们开始努力复兴物理学和生命的法则。一些哲学家将生命理解为一种使能量在生物体或生态系统边界附近局部转移的机制,而另一些人则将生命视为能量在未知潜力领域中的体现。生命对我们来说就像「盲人摸象」。生命能量及其转化的奥秘在整体上对人类来说还是未知的。像寓言中的盲人一样,我们试图根据自己对自然的有限经验和理解综合出一个连贯的画面。然而,理解生命能量、研究生命的法则,并希望预测生命的轨迹及其观测量,并非一个不切实际的目标。
我们中的许多人假装知道驱动生命的能量和功率。其实我们对能量的了解非常有限,尤其是关于生命和演化的知识。许多科学和人类社会长期存在的问题都可以直接与我们对能量及其转化的感知联系起来。安东尼·J·莱格特将任何学科中遇到的问题分为三个层次:
(i) 哈密顿量的,已知且可处理的;
(ii) 哈密顿量的,部分已知但难以处理的;
(iii) 哈密顿量的,甚至未知的。
系统的哈密顿量定义了系统的总能量。 我们可能知道与物质及其相互作用相关的哈密顿量,但我们不知道的是生命及其 演化 的哈密顿量 。理查德·费曼对科学家中的能量之谜持坦诚和认真的态度。他有一句名言:「重要的是要认识到,在当今物理学中,我们对能量是什么一无所知。」
基于20世纪的进展,我们可以从两种方式来定义能量:(a) 参照一个场 (引力场、电场、磁场) 或 (b) 参照一个频率f,如在公式 E=hf (其中h是普朗克常数) 所示。在基于经典场方法的科学中,我们隐含地假设物质的相互作用发生在无限的空间和时间领域中。然而,空间和时间并不是描述和研究能量的绝对要求。公式 E=hf 直接证明了能量超越了经典物理学的空间时间限制 (边界) 。因此,量子物理学应运而生。量子物理学通过考虑波函数来考虑能量的波动方面,并通过叠加、概率和各种类型的变换来研究相互作用。
采用经典物理学的场概念,我们之前提出了 社会场理论 (social field theory) 。社会场理论有助于量化与生命和生物体相关的能量。一个理论应该告诉我们测量什么。社会场理论建议根据社会力量(S)、个体力量(I)和一个信任向量(Γ)来测量与生命相关的能量。在经典场理论的框架下,我们用三个宏观变量定义了与生命相关的哈密顿量H:
H = SIΓ(在自然单位中)。(3.1)
能量是跨越物质、生命和演化的通用语言 。能量的时间导数,功率 (power) 在社会科学中的含义比经典热力学中更广泛。根据罗素的说法:「社会科学中的基本概念是功率 (power) ,就像能量是物理学中的基本概念一样。」社会场理论有助于定义与在社会领域中生命相关的能量。稍后,我们在纳维-斯托克斯方程的框架内,根据哈密顿量H和相关功率的定义,提出了一个关于能量的演化方程。
4. 物质
物质由基本粒子组成。物质占据空间,具有质量和惯性。惯性是物质在没有外力作用下继续保持静止状态或均匀运动的能力。在经典物理学中,物质是现实或可观测宇宙的基础,由空间和时间刻画。
物质科学在牛顿定律下形成。生命科学仍在发展中。要明确地划分物质和生命并不是一件容易的事情。病毒或处于休眠状态的种子是一些界限不太清晰的示例。从实际角度来看,我们根据积累的生命经验来区分生命与非生命。然而,科学要求我们在研究事物时,在物理量方面要更加精确。
5. 生命
生命是有组织的物质状态加上能量,它能够感知自身的能量状态和时间轨迹 。在某种程度上,生命可以被视为物质、电荷和信息,并与能量或功率相结合的各种循环的协同集合体。通过耦合,我们指的是一个循环产生的能量或功可以用来驱动另一个循环,以恢复其初始状态。因此,许多生命过程具有周期性:过程一次又一次地重复,并在各种空间和时间尺度上加强其他循环。显然,生命能够自我繁殖。
生命是一个能够感知自身存在、能量状态和演化的自主系统。这种对生命能量的隐含和主观感知将生命与非生命区分开来。生命容易体验却难以定义。关于生命的定义尚未达成共识。
像物质一样,生命占据空间并具有惯性。然而,生命具有内在的能量 (或功率) 来源,因此它可以自主地改变其状态。生命是一个开放系统,通过质量、能量和信息交换与周围环境相互作用。由于系统边界的存在,生命可以维持与环境不同的状态。
上面提到的许多关键词都是生命的特征。但特征并不总是足以定义一个主题,尤其是生命。我们避免用能量和功率来定义生命,因为这些概念仍存在一些模糊和不一致之处。生命的定义很容易产生更多的争议而不是清晰的理解。然而,上述任何情况都不应阻止我们研究生命和生命能量。
早些时候我们提到了物自在主义 (object egotism) 。物自在主义是牛顿物理学的一个基本概念。生命似乎拥有一种超越物自在主义属性的能力。生命可以改变一种反应,这可能超出了物质的能力。
因此,我们可以将牛顿第三定律 (方程 (2.1) ) 推广到生命领域,如公式 (5.1) 所示:R AB (t)=−β(Γ,τ) R BA (t±τ)。在这里,R是生命对外部刺激的一般反应,β是比例因子,τ是超前/滞后时间。例如,作为生物体的人类可以调节、超前或滞后反应力。这种反应取决于记忆、预见性等因素。
基于上述方程,我们可以区分物质和生命。可以说,物质遵循物自在主义进行反应,而生命体则可以对情况采取更全面的方法进行响应。根据这种解释,物质 (中的反应) 是瞬时发生的,而对于生命来说,响应可以在时间和空间上超前或滞后。
这项工作是对生命的一种尝试性阐释。然而,这种尝试部分受到了物理学传统方法的启发,这些方法旨在将自然现象追溯到简单的力学定律。这种对生命和非生命的划分可能会产生更多的问题而不是答案 。生命超越了工程和热力学启发的机器定义;生命随时间演化并能够自我繁殖。
6. 演化
在科学中,遵循牛顿第一定律,很自然地会从力 (内部或外部) 或精确的能量角度来思考演化。事实上,已经有许多尝试在物理学的框架内研究生命和演化。经过数百万年的演化,生物体会发生明显的变化。人类从猿类经过漫长的演化过程,表现出了明显的特征变化,如我们的祖先露西,据推测她是用两条腿行走的。在生物学中,演化是指随着时间的推移,基因物质发生变化的过程。 生物体与环境之间的持续相互作用,遵循类似于方程(5.1)总结的一般作用-反作用原理,可能导致基因改变、新性状出现,并最终形成新物种 。博弈论是社会科学中研究的类似作用-反作用现象。长期的相互作用可以改变生命、生物体以及环境的格局。生物体逐渐内化其基因中的变化,最终导致表型 ( 生物体的一组可观察特征) 发生变化。
在科普领域,我们归功于自然选择,它导致了更有可能生存和繁殖的生物体。自然选择是被提出解释生命和生物体演化的机制之一。合作和互惠也可能发挥重要作用。在种群中,自然选择过程可以增加有利等位基因的频率。科学要求用逻辑和物理量背后的法则来解释,比如动量、力、能量或功率。达尔文的自然选择理论中缺乏这种解释力。
生物体能够适应环境,同时能够影响和改变环境。由环境介导的变化源可以称为外部因素,可以视为表面力或外部功率。生命也拥有体力或内部功率。 我们推测生命的内部功率主要来自两个来源:(a) 作用-反作用不对称性,和(b)生命在其系统内部所蕴含的能量。 有时可能无法区分内部功率和外部 功率 。为了简化起见,我们可以将起源于生物体边界内的功率来源分类为内部功率,将与环境相关的功率分类为外部 功率 。生物体与环境处于持续相互作用中,就像鱼与水处于持续相互作用一样。很难确定一个水分子是鱼的一部分还是环境的一部分。

图1. (a) 物质在三维空间中的相互作用。(b) 生命单位在能量超空间 Ω(C1, C2, t) 中的相互作用。
图1通过对比物质在三维空间中的相互作用和生命单位在能量超空间中的相互作用,展示了生命现象的复杂性和抽象性。这种抽象描述有助于我们更深入地理解生命的本质和演化规律。
7. 演化之力
考虑两个或多个生命单位之间的相互作用。根据方程 (5.1) 中的广义作用-反作用关系, 两个生命单位之间的内力 R ij 不会相互抵消,并且通常不沿同一作用线 。 因此,生命单位系统总是存在一个合力以及相应的能量。 我们推测,这种生命所特有的合力是系统时间演化的原因。多体生命系统由于这种合力的作用而持续处于非平衡状态。
对于物质来说,如图1a所示,内力是共线的。力 R ij 和 R ji 在大小上相等但方向相反。因此,物质只对外部力作出反应。据此,牛顿的第一定律仅适用于物质,而不适用于一般生命体。
「让我们将生命单位的集合称为社会,Ω。在图1中,我们展示了物质 M 和生命 L 的集合,其中索引i、j和k分别代表每种类型的三个单位;G代表某种类型的质心,R代表反作用力。」
如图1a所示的物质,M i 和 M j 之间的内部力 R ij 和 R ji 相等且反向,并且具有相同的作用线。因此,它们的和 R ij + R ji = 0。然而,对于生命系统来说通常并非如此,如图1b所示。对于一个生命系统,R ij + R ji ≠ 0。在人类这样意识到自身能量状态的生物中,内部功率可能不总是平衡的。因此,一个多体生命集合Ω将在G点周围有一个净力和一个力矩。 我们认为这个净力/力矩及其相应的能量是 演化 的驱动因素 。
此外,生命系统还具有不同程度的内力R ii 。意识是生命单位感知环境以及自身的能力。人类可以利用意识,通过利用内力 R ii ,单独或与社会一起沿着能量阶梯上升。物质不能自行沿着能量阶梯上升。物质需要外部力来改变其静止或运动状态,这遵循牛顿的第一定律。但由于 R ii 和 R ij ̸=R ji 的存在,第一定律不适用于生命。达朗贝尔原理可能不适用于社会 Ω。换句话说,作用在生命体 L i 上的外部功率系统与这些生命体的有效功率系统并不等效。这可能是生命能够自发行动的原因,与物质的因果相互作用不同。 达朗贝尔原理通常适用于 R ij + R ji = 0 且不存在 R ii 的系统。
生命体如何调动其蕴含的能量来改变自身状态可能是显而易见的。在这里,我们关注的是作用-反作用不对称性如何导致合力。这种与生命相关的自主力可能是演化背后的物理原因。在非平衡态的物质中,我们也可能看到这种自主力。我们通常将这种力归因于熵产生的结果。
8. 生命演化的方程
我们可以遵循牛顿第二定律,用通常的符号写出物质的运动方程:
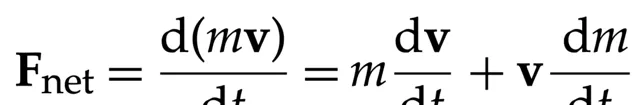 (8.1)
(8.1)
在假设质量为常量的情况下,上述方程简化为著名的 F net = ma。我们不知道生命是否能按照爱因斯坦的质能关系 E=mc² 将质量部分地转化为能量。然而,在本文所定义的框架中,生命能够感知其能量状态。空间和质量维度并不是生命的定义特征。因此,我们提出了一个关于生命演化的方程,该方程基于能量和功率。关于生命的功率方程形式如下:
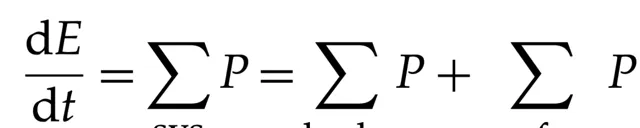 (8.2)
(8.2)
Stueckelberg 之前提出了一个类似的以功率交换来表述热力学定律的公式。在方程(8.2)中,我们以不同的方式对功率项进行了分组。
生命作为一个开放系统,由具有内部力量和功率源/汇的边界定义。在生命的基本单位中,线粒体可能体现了这种蕴含的能量源。我们可以区分体 功率 和表面 功率 。体 功率 是系统固有的,而表面 功率 则与边界和环境相关联。鱼可以使用体 功率 和表面 功率 来逆流而上或顺流而下。而死鱼则可能只受表面 功率 的作用而顺流漂下。这个例子生动地展示了生命系统如何通过内部和外部 功率 的相互作用来适应环境并维持其动态平衡。
方程右侧的功率项可以用生命体的力和类似速度的项来表示。这些力可以分为外在表面力(F ex )和内在体力(F en )。表面力主要来源于个体之间的相互作用 (即j ̸= i) ,如图1所示,而体力则是个体固有的。这些力可以计算为:
表面力:
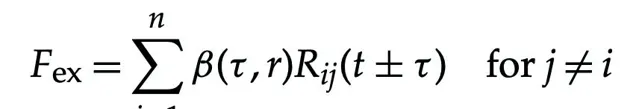
体力:F en = R ii (8.4)
一旦我们确定了与生命相关的力,就可以在纳维-斯托克斯方程的框架内写出演化方程:
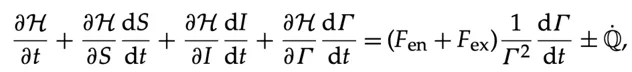 (8.5)
(8.5)
Q 的导数表示生命过程中固有的任何其他产生和耗散。
在本文中,我们定义生命为能够意识到能量状态。相应地,哈密顿量 H 可以用动能和势能等复合函数来表示。动能和势能的有序对定义了一个测量空间。对于生命的领域,我们将动能等同于个体的资本(C 1 ),势能等同于个体的能力(C 2 )。与经济学中的定义不同,资本和能力在社会场中都具有与能量单位等效的单位。因此,我们可以写
H = H(C 1 , C 2 , t) (8.6)
然后,生命演化方程也可以写成:
 (8.7)
(8.7)
方程(8.7)的第一项是哈密顿量 H 的局部导数,第二项和第三项是对流导数,右侧的项是源/功率项。
一个由N个生命单位组成的集合可以被称为社会S。我们可以定义社会(S)的哈密顿量(H s )为集合中所有哈密顿量的总和,即
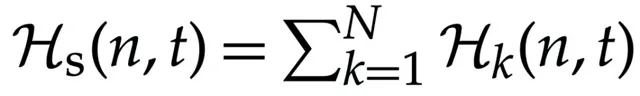
对于社会而言,方程(8.7)的形式类似于多变量 Fokker–Planck 方程,描述了绝对尺度下 H s 的时间演化。同时,在某些特定假设下,方程(8.7)也可以简化为 Lotka–Volterra 型方程,用于描述两种相互依赖物种的种群演化。
生命是多维的,因此描述生命的参数也可以是超维的。生命由各种物质循环、电荷循环以及由各种流动引起的和驱动的磁场循环组成。在生命的尽头,描述生命的个体强度(I)必须减少到我们用来描述物质的维度。如果我们承认这个参数I是多维的,包括物质、电荷、磁场 (包括电磁场和社会场) 等维度,那么我们可能能够在一个统一的框架内将物质和生命联系起来。
纳维-斯托克斯方程本质上是牛顿第二定律在流体流动中的扩展。根据设计,在纳维-斯托克斯方程框架内提出的功率方程可以简化为牛顿第二定律。在极限情况下,生命的法则和规则必须与物质的法则和规则相对应。这种对应性不仅体现了物理学原理的普遍性和一致性,也为我们理解和描述生命现象提供了重要的参考和依据。通过借鉴物理学的方法和理论,我们可以更深入地探索生命的本质和演化的机制。
社会场理论为我们提供了衡量社会中参数的思路。参数 S 和 I 是流动 (物质、能量、信息) 的函数,而信任向量 Γ 描述了流动和势能之间的关系。简单的维度分析表明,Γ 的单位等同于波长的倒数。这些是我们可以努力测量的参数,以便为物质、生命和演化的统一科学提供蓝图。
9. 结论
牛顿的三大定律为物质科学提供了结构。然而,这些定律对于研究生命和演化系统是不够的。本文展示了如何通过假设作用-反作用不对称性来实现对物质、生命和演化的综合理解。对牛顿第三定律在超越作用-反作用对称性的一般化,产生了一种可能与演化和驱动演化系统的动力相关的自主力。物质并不具备这种内在显著的力。生命是一个远离平衡的系统,已知具有内在的力和能动性。我们提出了一个用于研究生命的理论框架,在极限情况下对应于牛顿的物质定律。我们对生命科学的了解有限,然而,一个基于功率的框架似乎为我们关于生命和非生命的知识提供了一个统一的框架。我们提出了牛顿物质定律的一般化,可能成为我们在寻找物质、生命和演化统一科学过程中的指路明灯。












