前言
花生疮痂病是我国花生产区重要的真菌病害,产生光敏性真菌毒素痂囊腔菌,具有强烈的细胞毒性,为重要的毒力因子,前期研究发现,ESC生物合成极显著地受到光照调控,揭示病菌毒素生物合成的光调控作用及其分子机制,对探究该病菌的侵染过程、致病机制和调控网络具有重要意义,可为病害防治策略的高效制定提供了科学理论支撑。
本文开展了光质对病菌生长发育、ESC毒素产生及相关基因表达的影响效应研究,通过光质培养下样品进行了转录组测序及光受体基因挖掘,对花生疮痂病菌蓝光受体EaWC1基因克隆及生物信息学分析,构建了基因敲除载体并初步验证了蓝光受体EaWC!基因功能,研究结果旨在为花生疮痂病菌致病机制解析,探究光调控网络提供重要分子证据,为病害综合防治策略制定奠定科学的理论基础。

光质对真菌的调控作用
近年来研究表明,光质对真菌生长发育及次生代谢产物的影响极其显著,作为模式真菌粗糙脉孢菌的蓝光响应包括诱导孢子形成和性发育和菌丝体类胡萝卜素的合成。
与黑暗相比,白光和蓝光可以抑制轮枝镰孢菌)、早疫病菌和玉米灰病菌的分生孢子形成,促进生长和毒素含量产生,而黑暗和红光抑制构巢曲霉和竹黄菌分生孢子的产生,促进曲霉毒素和竹红菌素的产生。
黑暗时灰霉病菌对葡萄和番茄的致病力最强,白光及蓝光是致病力较弱,反而绿光对其在葡萄和草莓上致病力具有很强的抑制作用,不同光质下培养5株不同的性状的蛹虫草菌株,均表现为蓝光抑制其菌落生长,白光和蓝光极显著促进分生孢子的产生。
黑暗条件下仅产生少量分生孢子,真菌光质响应的研究揭示了一种意想不到的复杂性,不同物种间对光质响应存在很大差异。
蓝光受体
真菌对450nm(蓝色)至700nm范围内波长的光有光响应,蓝光受体更易受光刺激产生光信号,快速启动光感系统,调控下游次生代谢产物编码基因表达而备受关注,目前已经在真菌中鉴定了几类蓝光受体,包括WCI,WC2,VIVID,隐花色素以及光解酶。
粗糙脉孢菌中真菌光受体的分子机制已在得到了较为详细的研究,最早从粗糙脉孢菌中克隆得到第一个真菌蓝光受体基因WCI,该受体对N.crassa昼夜节律、生长趋向性、无性发育、有性生殖、抗UV损伤和类胡萝卜素等具有调控作用。
目前在构巢曲霉、烟曲霉、禾谷镰刀菌、卷枝毛霉菌、深绿木霉、交链格孢、灰霉菌和玉米灰斑病菌等多种真菌中均筛选鉴定到蓝光受体ⅣC1同源基因。
真菌光受体在结构上拥有多功能结构域,蓝光受体WC1编码蛋白通常具有三个PAS结构域和一个锌指结构域,其中两个PAS域用于蛋白质-蛋白质互作,锌指结构域用于核定位,靠近N端的PAS域属于一类特殊的结构域,称为LOV结构域,也称光、氧或电压域。
之后不久克降了蓝光检测所必需的第二种蛋白质,命名为WC2,具有一个锌指结构域和单个PAS结构域,可定位于细胞核,WC蛋白对于真菌光响应具有调节周期节律,促进或抑制有性发育,分生孢子产生及次生代谢产物合成积累的功能。
VIVID是一种小蛋白质简称VVD,具有一个与FAD或FMN结合形成的LOV结构域,在光照后基因表达的瞬间激活中起重要作用,与WC1和WC2不同的是,在脉孢菌核中没有发现VVD,只有在光诱导后才能在细胞质中观察到活性物质,VIVID对生物钟启动极为重要,光信号诱导生物钟启动会调节生物内部从而适应生长环境。
接收蓝光到蓝紫波段光信号的受体还有隐花色素,它在原核和真核生物中均被发现,其广泛的存在于构巢曲霉、粗糙脉孢菌、玉米灰斑病菌、木霉和核盘菌等多种真菌中,其重要调控真菌的光修复,生物钟,有性无性发育和次生代谢等。
光敏色素最先是在植物中发现的,人们对其进行了充分的研究,在真菌中,这种光感受需类型首先在两种模型真菌构巢曲霉和粗糙脉孢菌中被发现,目前关于真菌红光受体的研究还主要集中于曲霉中,其他真菌中也被发现如灰霉岗。
光敏色素是一种大型的蛋白质,由一个结合发色团的光感区和一个信号输出域组成,感觉区域由三个区域组成域,GAF小配体结合区域和PHY光敏色素域。
面信号输出区域与原核组氨酸激相似,包括HKD组氨酸激酶结构域,HATPase腺苷三磷酸酶结构域,RRD响应调控域,光敏色素编码基因Fph同样调控真菌重要的生理生化过程,目前的研究主要集中在有性无性发育和色素沉着上。

尽管已经证明构巢曲霉中FphA可以使用胆绿素,但真菌光色素中胆红素发色团衍生物的性质仍有待确定,发色团与PAS或GAF域结合并存在于两种可互换的构象中,它们的光谱特性很好地区分开案,光敏色素以P形式吸收红光,从面改变构象并将吸收最大值移至远红光光谱,这个新形式叫做p,在吸收远红光时,发色团恢复到Pr形式完成光响应循环。
蓝光受体对真菌次生代谢的调控机制
细胞生物中光感应与传导是一个复杂的生物过程,作为对真菌影响最广泛显著的蓝光,蓝光受体是各种真菌生长发育和次生代谢等生理生化过程所必须的生物感受器,众多学者对蓝光受体功能进行研究。
光生物学研究表明,蓝光受体WCI基因的表达调控与老种真岗次生代谢产物的生物合成具有高度关联性,光照能够促进水积恶苗病菌中镜孢菌素的生物合成,此时蓝光受体co4基因高表达,模式真菌粗糙脉孢菌中C-1基因影响生物钟和类胡萝卜素的生物合成。
禾谷镶抱菌中蓝光受体Fgwcl基因却主要影响有性繁殖和无性发育,灰霉菌中红光受体光敏色素Bqphy2,Bcphy3基因的缺失均抑制黑色素的生物合成,但突变株的致病性却不相同,在缺失2基因的突变体中致病力显著增强,面缺失Bcpp基因的突变株致病力明显降低。
链格抱菌中蓝光受体De基因强烈诱导了毒素的生物合成,但负调节Alternariol(AOH)毒素产生,玉米尾孢菌中蓝光受体基因CRPI表现出促进毒力因子尾孢菌素积累,CRPI基因缺失明显降低病菌致病性和病斑扩展能力。
雪洲镜孢菌的研究中发现,蓝光受体FamC!在感知光信号、调节抗紫外线(UV)损伤、次生代谢、有性生殖及致病力中同样是必需的,该基因的缺失导致光诱导的光解酶基因表达的缺失,从面导致对紫外线的过敏。
蓝光受体用C1在真菌生长发育、次生代谢和环境感知中所行使功能具有复杂性和多样性,调控模式和机理亦相差甚远,1.3.2细胞中其它光信号转换。
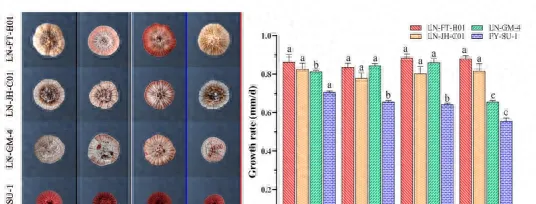
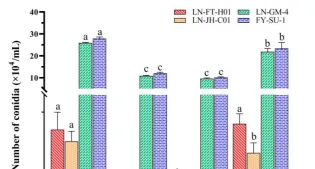
菌株培养于PDA平板上,置于25℃光照培养,培养时间分别为14d、21d、28d.35d和42d,随后打取菌饼,烘干后以丙酮为提取溶剂萃取,采用超声振荡法提取至无色后合并,减压浓缩后为病原菌毒素粗提液。
定量检测参考刘璐(刘璐等,2018)方法,以紫外分光光度法绘制标准曲线,采用标准曲线法定量毒素,选择最适培养天数,进行后续毒素测定的试验。
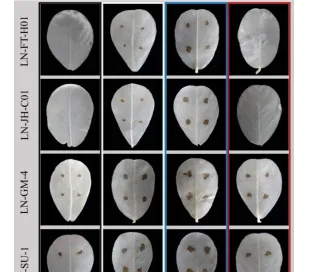
采用菌悬浮液定点定量离体接种法接种菌株,根据2.1.2制备菌悬液,取苗后14-28d健康无病,长势均匀,展开的花生叶片,用无菌水轻拭叶片,对花生离体叶片进行保湿培养。
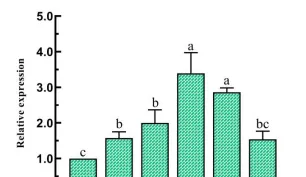
在叶片设置4-6个对称的接种点,以无菌水为对照,每接种点接种20L菌悬液,置于不同光质条件下,22-25℃保湿培养,每日观察发病情况,接种15d时拍照,脱色记录病情。
差异表达基因分析
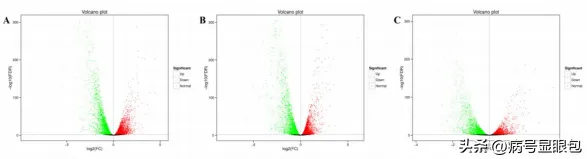
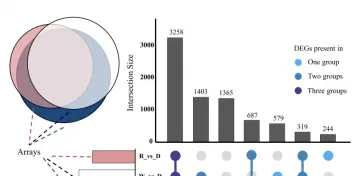
以同条件下黑暗处理的花生疮痂病菌作对照,分析不同光质条件处理36h后的差异表达基因,WVSD共有5926个差异基因,其中2877个基因上调,3049个基因下调:BvsD共筛选出6344个差异基因,其中3096个基因上调表达,3248个基因下调表达:RvSD共有4507个差异表达基因,其中2094个基因上调,2413个基因下调。
说明本研究中不同光质处理下花生疮痂病菌的基因表达受到调控,处理中差异表达基因数量关系,与黑暗相比,只在蓝光处理时,显示的差异表达基因有1365个,只在白光处理时,显示的差异表达基因有579个,只在红光处理时显示的差异表达基因有244个。
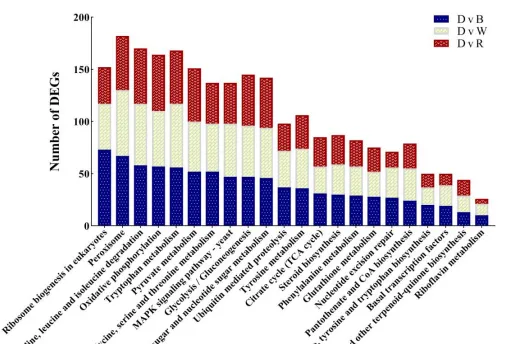
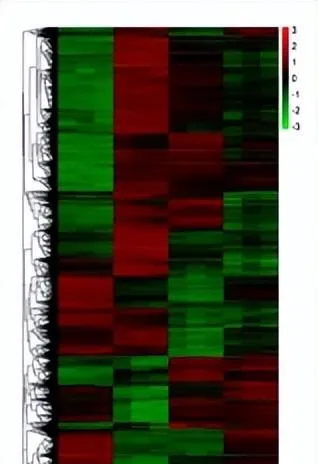
白光和蓝光处理下差异表达基因远远大于红光处理时数量,表明与黑暗相比,白光和蓝光处理诱导基因的表达比红光处理时多,蓝光条件处理下诱导的基因表达最多,差异表达基因聚类分析图中也存在相似的趋势,差异表达基因上调倍数越大,颜色越红,反之越绿。
黑暗处理时上调的基因,在白光和蓝光处理时下调黑暗处理下调的基因,在白光和蓝光处理时上调,红光(R)处理时基因表达与黑暗基因一致,这表明白光和蓝光处理与黑暗对照处理之间存在很大的差异,在红光处理时的基因表达和黑暗处理相似。
结论
为探究光对于花生疮痂病菌基因表达的调控,共筛选到474个受光质诱导表达的基因,包括11个假定光受体基因,57个转录因子,81个参与光信号通路基因,352个与次生代谢相关基因,通过Blast比对及系统发育树分析表明,EVM0003940为WC-1同源基因,EVM0001480,EVM0004523,EVM0002126和EVM0003727均为隐花色素同源基因。
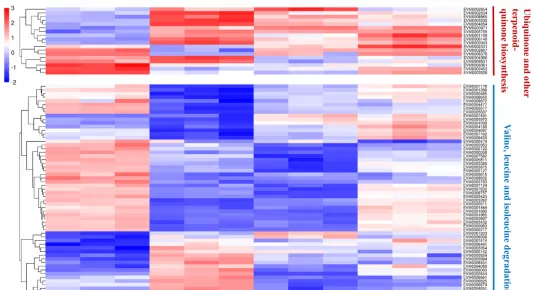
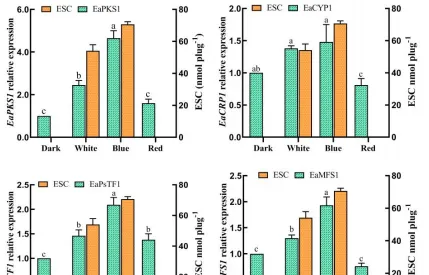
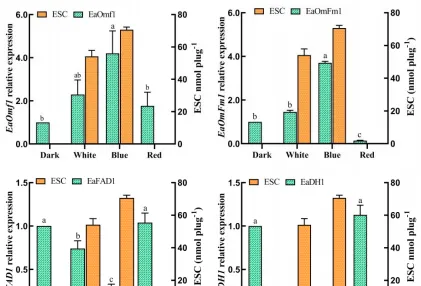
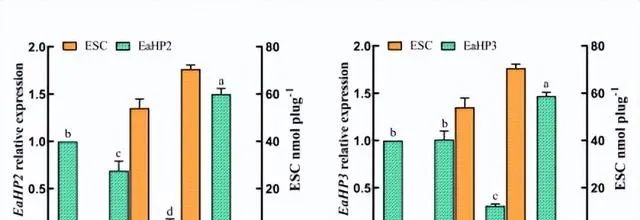
不同光质条件对花生疮痂病菌ESC毒素产生具有显著影响,对代谢途径中转录组数据的DEGs分析有助于识别花生疮痂病菌花醌类化合物通路中基因在不同光质处理下的表达模式,通过Blast对不同光质处理下的DEGs进行功能分析。
预测325个可能是参与次生代谢产物生物合成的基因,根据这些基因的功能注释,主要集中在9个功能组,分别为聚酮合酶、非核糖体肽合成酶、FAD依赖性单加氣酶、甲基转移酶、FAD依赖性氣化还原酶、NADPH脱氢酶、转运蛋白、细胞色素P450和铁还原酶类蛋白。
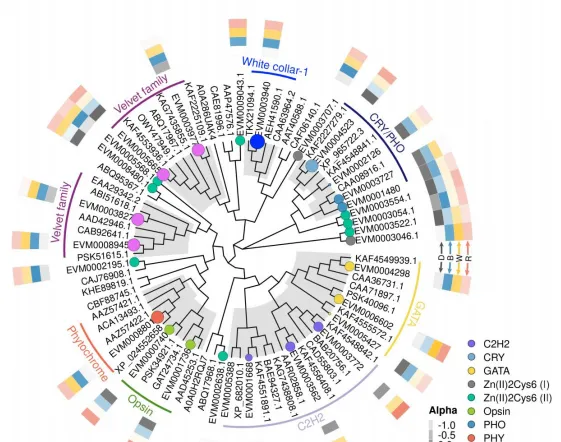
通过Blast和系统发育分析,成功筛选出12个次生代谢相关基因,通过荧光定量PCR表达验证,结果表明,这些基因的表达模式与转录组表达模式基本一致,这些基因调控参与了痂囊腔菌素生物合成,为进一步分析提供了研究基础。











