

次級代謝產物是以微生物、植物或動物等細胞合成的初級代謝產物為原料或前體,透過一些特殊代謝途徑合成的一類可發揮顯著生物活性的代謝產物。這類代謝產物大多具有特殊的生理活性,在機體適應特殊生境或完成特定代謝的過程中具有重要作用,也是中草藥和天然藥物的重要藥效物質,因此挖掘此類代謝產物已成為尋找具有潛在天然活性產物的主要手段之一[1-2]。
放線菌分布廣泛,因其可產生大量結構新穎且具有豐富活性的次級代謝產物而備受關註,包括抗生素、免疫抑制劑、抗腫瘤藥物、維生素、酶等[3]。鏈黴菌 Streptomyces 屬於經典的放線菌,其產生的次級代謝產物一直都是抗生素類藥物發現和開發的重要來源。截至目前,在醫藥、農業等領域使用的抗生素中有90%均為放線菌合成的次級代謝產物,如大環內酯類抗生素、胺基糖苷類抗生素、四環素、醯胺醇等,而50%以上的鏈黴菌都能產生抗生素[4-5]。
盡管如此,在自然生長狀態下,野生鏈黴菌的次級代謝產物合成效率較低,遠不能滿足大規模生產需要,其原因主要有:鏈黴菌是一種絲狀細菌,具有復雜的發育周期,而其次級代謝產物的合成與鏈黴菌生命周期中的形態分化密不可分[6];鏈黴菌次級代謝產物的生物合成基因簇(BGCs)在自然生長條件下大多處於沈默或低水平表達狀態[7],限制了鏈黴菌次級代謝產物種類和產量的積累;
此外,目前關於鏈黴菌次級代謝產物合成的代謝網絡調控相關機制仍未得到充分解析,但隨著全基因組測序技術和合成生物學技術的快速發展,大量的鏈黴菌BGCs被挖掘,如何啟用這些BGCs提高鏈黴菌次級代謝產物的產量已成為新藥研發領域尋找具有顯著活性化合物的研究熱點[8-9]。基於此,本文對調控鏈黴菌次級代謝產物生物合成的非靶向和靶向調控策略進行綜述,為實作鏈黴菌次級代謝產物生物合成效率的高效強化提供參考。
1 鏈黴菌發育周期及其次級代謝產物的產生
鏈黴菌是一類廣泛存在的革蘭陽性絲狀原核微生物,呈現出復雜的發育周期和生長模式。鏈黴菌生命周期始於分生孢子的萌發,當分生孢子在適宜條件下吸水後透過伸長和分裂啟動發芽並以放射狀向基質內層和表面擴散,形成大量具有吸收營養和排泄廢物功能的基內菌絲體,同時又不斷向上分化出氣生菌絲。該階段鏈黴菌菌絲處於快速生長狀態,當氣生菌絲成熟後,鏈黴菌會進入孢子分化階段,分化成孢子絲,而後透過橫割分裂的方式,產生分生孢子。孢子分化是鏈黴菌生命周期中的一個重要轉變過程,該過程通常伴隨著菌絲形態的變化和次級代謝產物的合成[5,10]。
液態發酵是鏈黴菌規模化培養進行次級代謝產物生產的主要方式,且通常將發酵過程分為菌體生長和產物合成2個階段。以抗生素為例,其通常在菌體生長階段結束後由基質菌絲體產生,目前在液態發酵環境中鏈黴菌菌絲分化領域的研究工作主要集中在菌絲形態、培養基組成和生物反應器設計等[11]。鏈黴菌在液態發酵體系呈現的形態可分為4種類別:小球(直徑950 μm的致密團塊)、團塊(直徑600 μm的不太致密團塊)、分支菌絲和非分支菌絲,部份研究者認為,細胞聚集形成小球和團塊是次級代謝產物高效合成的基礎[12-13]。因此,鏈黴菌次級代謝產物的合成與其形態分化過程密切相關。
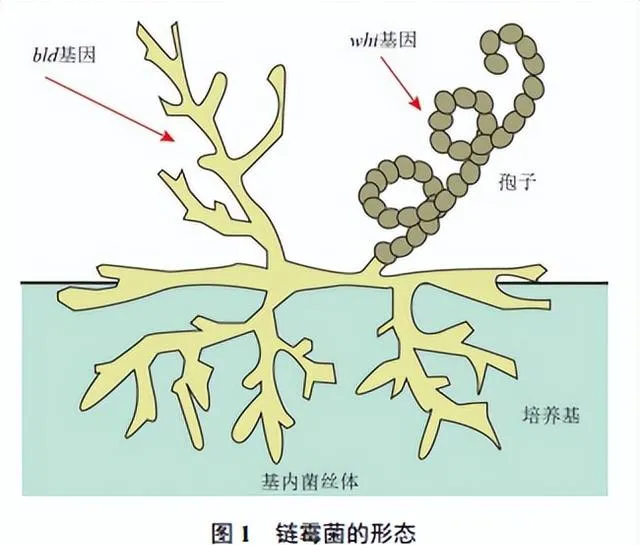
鏈黴菌的形態分化是多個調控因子和調控網絡綜合作用的結果,其中 bld ( bald )家族和 whi ( white )家族基因發揮著關鍵的調控效應(圖1)[14]。 bld 家族基因包括 bldA、bldD、bldM 和 bldN 等,其中 bldA 基因能編碼一種特殊的tRNA分子,透過轉譯UUA(TTA)密碼子而影響蛋白質合成; bldD 基因參與抑制鏈黴菌氣生菌絲發育和孢子形成相關基因的轉錄,阻止孢子在菌絲生長期的生成; bldM 基因則編碼一種雙組分系統孤立應答調控蛋白,參與細胞的應答和調控; bldN 基因編碼的是一個胞質外功能σ因子,也是一種轉錄調控蛋白。可見, bld 基因家族編碼一系列的轉錄因子,透過調控鏈黴菌形態分化過程中的細胞合並、細胞壁合成與分化等關鍵步驟而對鏈黴菌的代謝發揮調控效應,這些基因的突變或缺失可導致鏈黴菌的分化受阻[14-15]。
whi 家族基因在鏈黴菌的形態分化和孢子形成過程中也發揮著重要調控作用,其中, whiA 和 whiB 表達產物是參與細胞延伸和分裂時期轉換的關鍵調控因子,能夠影響細胞分裂和孢子分隔的形成; whiG 基因編碼一種RNA聚合酶σ因子,主要在鏈黴菌的氣生菌絲分化晚期發揮作用,調控進入孢子生成階段; whiI 基因編碼雙組分系統孤立應答調控蛋白,參與細胞的應答和適應力變化[14,16]。
在鏈黴菌的形態分化過程中,這些基因和調控因子透過調控基因表現、訊號傳導和細胞功能,共同參與調控細胞的合並、細胞壁合成、菌絲分化和孢子形成等關鍵步驟,它們的相互作用和調控網絡對於鏈黴菌的生長發育、代謝產物合成及適應力變化等具有重要影響,對於鏈黴菌資源的研究和開發具有重要參考價值[14-17]。鏈黴菌形態分化的周期調控是一個復雜過程,涉及多個基因和調控因子的相互作用和調控網絡。
目前,鏈黴菌的分化過程和生命周期已被深入研究,但從形態分化到次級代謝產生的轉變機制仍不清楚,一旦闡明這些機制,或特許以透過調控某些節點開關對次生代謝做出正向調節,進而實作高活性次級代謝產物的強化合成,因此鏈黴菌中形態分化關鍵基因的表達研究對闡明次級代謝產物合成相關的代謝通路和最佳化鏈黴菌生命周期具有重要意義。
2 鏈黴菌的代謝調控策略
大量的鏈黴菌基因組測序結果表明鏈黴菌含有豐富的調控次級代謝產物合成的BGCs,但透過生物資訊學分析發現BGCs中的很多基因在常規培養條件下都處於沈默狀態[18],從而使鏈黴菌獨特的次級代謝產物合成途徑處於關閉或低效率代謝狀態。因此,啟用和調節鏈黴菌沈默的BGCs以促進次級代謝產物種類增加和產量提升是鏈黴菌源高活性化合物開發的研究重點[19]。
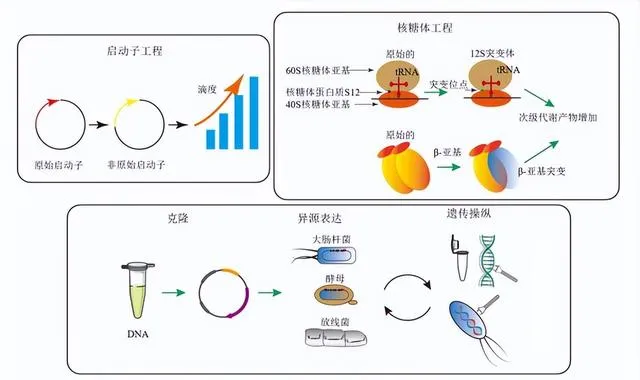
目前,已有多種啟用BGCs的方法得到套用,包括非靶向啟用(細胞外訊號誘導,圖2)和靶向啟用策略(重建生物合成途徑和關鍵蛋白異源表達,圖3)[20]。值得註意的是,非靶向調控策略可能不具有高度選擇性,而且對於產物產量和結構的影響往往也具有不穩定性,因此,在套用非靶向調控策略時,需進行調控策略和培養條件的最佳化,並結合分子生物學和代謝組學等方法深入研究其機制,進而實作對鏈黴菌生長和次級代謝的理性調控;而靶向調控策略結合了基因工程、合成生物學、訊號通路研究和代謝工程等方法,可以更精確地調控鏈黴菌次級代謝產物的合成,對於鏈黴菌源的成藥性化合物開發具有重要套用價值[18-21]。

2.1 非靶向調控策略
2.1.1 培養基組分的調整
培養基組分對鏈黴菌次級代謝產物的合成具有重要影響,不同培養基組分的組合和濃度可以影響菌株的生長和代謝途徑的調控。大量研究表明,不同培養基營養條件(碳源、氮源、能源、無機鹽、水)組成對鏈黴菌的次級代謝產物的種類和產量有明顯影響[22-23],碳、氮源可提供進行能量代謝和蛋白質合成的必要物質基礎,而無機鹽和微量元素則可影響鏈黴菌代謝反應的速率和方向[24-25]。
微生物的能量代謝涉及電子傳遞過程,微量元素如鐵(Fe2+/Fe3+)和鉬(Mo2+/Mo4+)參與細胞色素和鐵硫簇的形成,參與電子傳遞鏈傳遞,同時鐵作為細胞代謝中大量關鍵酶的輔助因子,是重要的必需金屬離子之一,但是鐵在非酸性的好氧培養環境並不容易獲得,鏈黴菌以其產生各種鐵載體的能力而聞名,缺乏鐵載體產生或攝取能力的鏈黴菌菌株生長可能會減少甚至消除,從而導致其形態分化和次生代謝物的合成受阻,而鐵載體的生物合成與培養基中的鐵含量密不可分[26]。
此外,鋅(Zn2+)和鎘(Cd2+)也是大量酶的輔助因子,錳(Mn2+)在一些氧化還原酶中發揮重要作用。鋅已被證實具有在細胞中發揮穩定蛋白質結構折疊和作為酶催化輔助因子的雙重作用[27],但高濃度的Zn2+也會對微生物的生長代謝發揮負效應,因此,必須嚴格調節和維持鋅穩態[28]。銅(Cu2+)作為一種還原性金屬離子,雖然對細胞有潛在危險,但銅(Cu2+)參與酵母色素和氧化還原酶的形成,從而促進電子傳遞和催化還原反應,特別是在鏈黴菌中,培養基中銅含量對形態分化的開始和次級代謝物的產生具有顯著影響[29]。李全樂[30]透過考察Fe2+和Mg2+對吸水鏈黴菌 S. hygroscopicus 代謝組的影響,發現Fe2+在一定程度上可影響吸水鏈黴菌胞內脂肪酸和胺基酸合成並促進雷帕黴素的合成,而Mg2+對吸水鏈黴菌胺基酸代謝影響也比較顯著。
此外,生長因子也是微生物生長和代謝所必需的重要成分,如維生素、脂肪酸、胺基酸等,一些些微生物需要外源的生長因子才能正常生長;調節劑和激素也可以調控微生物的生理過程和代謝途徑。如去氧膽酸可作為調節劑,影響某些細菌的次級代謝產物合成,而培養基中添加抗生素可以抑制其他微生物的生長,從而實作目標微生物的選擇性培養,一些抑制劑也可用來阻斷某些特定代謝途徑或抑制細胞特定酶活性[25]。此外,部份強氧化劑也會對其有較大的影響。Wei等[31]發現最佳化吸水鏈黴菌5008培養體系中pO2的添加量和添加時間,可以將纈黴素A的產量提升40%。
2.1.2 培養環境的改變
微生物培養過程中環境溫度、pH、溶氧量、滲透壓、容器類別等也會引起其相關基因的差異表達[32],次級代謝相關途徑中關鍵酶的活性直接影響次級代謝產物合成,如培養溫度過高導致酶失活,溫度過低則影響酶促反應速率,同時滲透壓、pH、溶氧量等也會引起微生物生化反應速率的變化,並對一些沈默BGCs的啟用發揮調控效應[33]。Boukaew等[34]透過對比不同的溫度、光照/黑暗條件下鏈黴菌 S. philanthi RL-1-178、 S. philanthi RM-1-138、 S. mycarofaciens SS-2-243的代謝狀況,發現不同溫度和光照/黑暗條件對RM-1-138菌株合成的抗真菌代謝產物種類和產量影響極為顯著,而Zhang等[35]發現透過控制灰略紅鏈黴菌 S. griseorubens JSD-1發酵過程中的pH變化和溶氧量,可使該菌株纖維素酶、木聚糖酶和濾紙酶的產量大幅提升。
2.1.3 微生物共培養
微生物共培養是指2種及以上微生物(包括細菌、絲狀真菌、酵母等)在同一培養體系中共同生長和相互作用的過程。該過程可實作不同菌株間代謝產物和代謝訊號的共享和交換,進而發揮不同菌株間的協同或拮抗作用,啟用可能的沈默BGCs,實作微生物新代謝產物的合成,或目標代謝物產量的顯著提升[36]。
微生物共培養可以產生多種效應,如共生、協作、競爭等。伴侶關係是指微生物之間透過相互利用代謝產物或提供生長因子等互相促進生長;協同作用是指不同微生物的共培養可能導致協同效應,即彼此之間生長和代謝的相互促進,從而提高某些次級代謝產物的產量;競爭關系是指共培養體系中不同微生物之間爭奪有限的可利用資源,包括營養物質和生長空間,進而導致對一種或多種微生物的生長抑制或排斥,競爭關系的出現取決於不同種微生物之間的生態席位差異和生長速率等因素。
共培養體系中互為競爭關系的微生物可透過產生抗生素來抑制競爭對手的生長。這種抗生素產生可以被視為一種防禦機制,幫助微生物在共存環境中獲得有利競爭。在共培養中,微生物之間還可以進行基因水平的相互作用,如基因轉移、共享或調控等。這些相互作用可以導致基因型的改變和適應力特征的轉移[37-40]。微生物共培養方法已被大量研究證實有效,如Shin等[41]將海洋鏈黴菌與芽孢桿菌共培養產生一種新的含哌酸環肽;Huynh等[42]將鏈黴菌 S. sp. GA02和 Pandoraea sp. GA02N共培養產生了2種芳香化合物。
綜上,微生物共培養技術現已經被廣泛用於合成生物學領域,相比於單一菌株培養,微生物共培養不僅可以減輕出發菌株的代謝負擔,實作復混成合物的協同合成,還可以充分發揮不同物種的優勢和能力,利用低劣生物質以提高目標產品經濟性[43],但目前還存在共培養體系穩定性差、自主調控能力缺乏、關鍵控制點不清晰、工業放大難等問題。
2.1.4 中藥萃取物調控
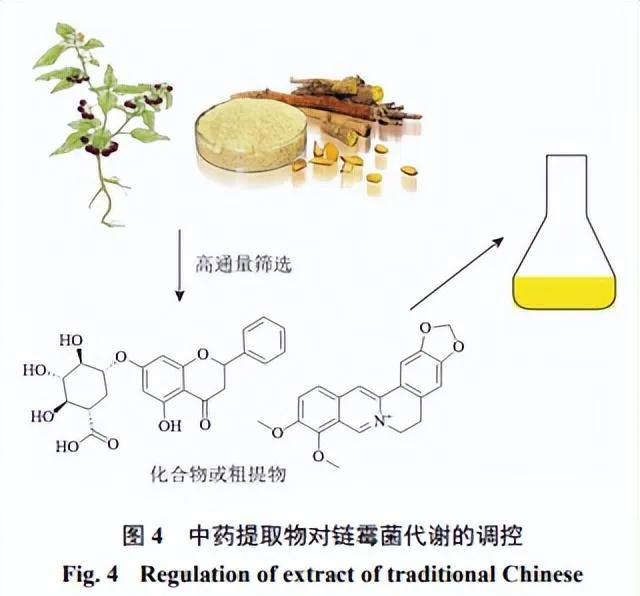
中藥萃取物中的活性成分,如生物堿、黃酮等具有抑菌或抗微生物活性[44],這些活性成分可以透過抑制微生物的代謝酶活性、破壞細胞膜結構或抑制細胞分裂等,抑制微生物的生長和繁殖(圖4)[45-46]。一些中藥萃取物中含有具有激素作用的成分(植物生長素、植物激素等),這些激素可以刺激微生物的細胞分裂、營養吸收和代謝活動,加速其生長和繁殖過程[47]。酚類化合物可以改變微生物膜結構,從而影響細胞透過性和物質轉運過程,進而影響微生物的代謝效率和產物合成[48]。
抗氧化、抗炎成分可以增強微生物的抗逆能力,這些抗逆能力的提高可以使微生物在惡劣環境下更好地適應和生長[49]。Seyedsayamdost[50]於2014年概述了一種利用高通量篩選小分子化合物啟用沈默BGCs的方法。這種小分子透過小分子文庫的高通量篩選提供了潛在的誘導劑,而中藥是天然活性分子化合物的寶庫,透過高通量篩選中藥中某些具有較強生物活性的小分子可以作為激發子啟用沈默的BGCs,雖然目前發現的這種激發子大多是抗生素,
但聶智英[51]將鏈黴菌CB02414、CB03234、CB02130和NRRL S-1813培養液中加入杜仲葉水提物後透過液質聯用技術分析發現,加入杜仲葉水提物的CB02414培養組比對照組新增了2個化合物,加入杜仲葉水提物的CB03234培養組比對照組新增了1個化合物,且該化合物的產量隨著杜仲葉水提物濃度的增加而增加,同時發酵液粗提物抑制金黃色葡萄球菌 Staphylococcus aureus 的活性也得到了明顯增強,加入杜仲葉水提物的NRRL S-1813培養組比對照組新增了1個化合物,同時發酵液粗提物對金黃色葡萄球菌和藤黃微球菌 Micrococcus luteus 的抑制效果也顯著增強。上述數據提示中藥中的某些小分子可作為激發鏈黴菌中沈默BGCs的激發子,為沈默BGCs的啟用提供了一種新的策略,但其具體機制仍需進一步研究。
2.1.5 表觀遺傳修飾
表觀遺傳修飾可以在不涉及DNA序列改變的前提下對基因的表達進行調控,通常透過調控DNA甲基化、組織蛋白修飾和非編碼RNA改變基因的表達水平或沈默狀態(圖5)[52]。DNA甲基化指在DNA甲基化轉移酶的作用下,基因組CpG位點(C和G相鄰的堿基對)二核苷酸的胞嘧啶5號碳位共價鍵結合1個甲基基團,DNA甲基化的結果通常導致基因沈默,抑制基因的轉錄活性[53]。而組織蛋白修飾是一類常見的表觀遺傳修飾,一般透過化學修飾改變組織蛋白分子狀態,從而影響染色質的結構和功能,達到調節基因的可及性和轉錄活性的目的。
常見的組織蛋白修飾類別包括以下幾點:
(1)乙酰化:組織蛋白乙酰化通常與基因的活性相關,透過加上乙酰基基團,可以使染色質更松弛,使得轉錄因子和轉錄機器更容易接近基因組,促進基因的轉錄;
(2)甲基化:組織蛋白甲基化是在賴胺酸殘基上添加甲基基團,可以引起不同的效應,在一些位點,組織蛋白甲基化可以導致染色質結構的緊縮,抑制基因轉錄;
(3)磷酸化:組織蛋白磷酸化通常與活化基因表現相關,磷酸基團的加入可以導致染色質結構的改變,促進轉錄因子與染色質的相互作用,並啟用基因轉錄;
(4)泛素化:組織蛋白泛素化是在賴胺酸殘基上加上泛素分子,它可以參與染色質結構的調節和蛋白質的降解,並影響基因的表達。
此外,還存在其他類別的組織蛋白修飾,如絲胺酸磷酸化、絲胺酸/蘇胺酸乙酰化、絲胺酸/蘇胺酸甲基化等,這些組織蛋白修飾可以透過與其他轉錄調控因子的相互作用,調節染色質的結構及轉錄的啟動和終止過程[54]。

組織蛋白去乙酰化酶和甲基轉移酶常作為化學表觀遺傳修飾劑啟用真菌的沈默BGCs,這是因為真菌DNA結合在組織蛋白上,組織蛋白將DNA壓縮成染色質並調節復制和轉錄[55],且這些基因的位置往往靠近其染色體的端粒,而組織蛋白乙酰化和DNA甲基化對這些位點的轉錄有很大影響,可抑制或關閉真菌次級代謝產物的生物合成。
鏈黴菌等原核生物染色體結構雖比真菌染色體結構簡單,但化學表觀遺傳修飾劑也可對鏈黴菌的次級代謝產物發揮調控效應[56],如Kumar等[57]用5-氮雜胞苷25 μmol/L處理 S. coelicolor AZRA 37後,液相色譜分析表明發酵產物增多,抗菌活性增強,全蛋白分析表明出現了新的孔蛋白。可能是由於鏈黴菌的DNA、蛋白質和RNA構成的染色體是高度壓縮的超螺旋結構,DNA環繞在由RNA和蛋白質形成的「擬核」的內外,一些基因簇也可能由於壓縮導致沈默。此外一些啟動子附近DNA的甲基化也會影響基因表現,使用DNA甲基化酶抑制劑抑制其DNA甲基化,可啟用相關基因表現[57-58]。
2.1.6 訊號分子蛋白的表達調控
訊號分子蛋白的表達調控是細胞中的一種重要機制,可以全域調控基因表現水平以響應特定的環境訊號或細胞狀態。目前,在鏈黴菌中報道訊號分子合酶只有2種[59],包括A因子合成蛋白(AfsA)和乙酰輔酶A氧化酶(Aco)同源蛋白。AfsA同源蛋白可催化脂環類化合物的合成,如γ-丁內酯、丁烯羥酸內酯或呋喃類訊號分子;Aco同源蛋白主要催化丁烯羥酸內酯類訊號分子合成的酶[60-61]。
在鏈黴菌中,AfsA同源蛋白可透過催化特定的化學反應生成γ-丁內酯、丁烯羥酸內酯或呋喃類訊號分子,這些訊號分子在鏈黴菌的生命周期和發育過程中具有重要的調控作用,丁烯羥酸內酯類訊號分子還可影響鏈黴菌細胞分化、發育和次級代謝產物的合成。AfsA和Aco同源蛋白的表達和活性均受到環境條件和細胞狀態的調控,可作為鏈黴菌次級代謝調控的重要訊號分子蛋白[62-63]。但是目前有關Aco類訊號分子合酶的功能認知仍需進一步深入探索[59]。
此外,全域調控蛋白在調節鏈黴菌的初級和次級代謝轉換過程中也發揮著關鍵作用[64]。如全域調控蛋白GlnR[65]和PhoP[66]分別透過監控鏈黴菌的氮代謝和磷酸鹽代謝狀態來控制抗生素的生物合成。研究表明全域調控轉錄阻抑蛋白DasR是一個平衡鏈黴菌碳源和氮源代謝的關鍵交叉點,它透過與中間代謝產物6-磷酸乙酰胺基葡萄糖(GlcNAc-6P)和6-磷酸胺基葡萄糖(GlcN-6P)結合,參與了鏈黴菌的營養感應、形態分化、初級代謝和次級代謝轉換等生物學過程[67]。
N -乙酰葡糖胺的代謝產物GlcNAc-6P和GlcN-6P與DasR蛋白結合後,透過調節蛋白的構象,解除DasR與響應元件( dre )位點對下遊基因的抑制作用。表明GlcNAc-6P和GlcN-6P透過改變DasR蛋白的構象來調節其與 dre 位點的結合,從而減弱甚至消除對下遊基因的阻礙作用[68]。DasR蛋白的研究對於了解細菌的基因表現調控網絡和代謝途徑具有重要意義,為新抗生素等次級代謝產物挖掘和微生物代謝工程技術創新提供依據,然而,還有許多關於DasR蛋白的具體機制和功能目前仍未得到充分解析[69]。
2.1.7 轉錄因子調控
轉錄因子是一類可以與DNA分子上的特定核酸序列結合,並對基因轉錄發揮調控效應的蛋白質,其透過與DNA結合而參與調控基因轉錄起始復合物的形成或抑制,從而控制基因的轉錄水平糊轉錄速率[70]。在鏈黴菌中,轉錄因子調控主要透過調控轉錄因子基因簇和第2信使系統等方式實作。鏈黴菌的基因組中通常存在一些特定的轉錄因子基因簇,它們編碼具有相似結構和功能的轉錄因子,這些轉錄因子能夠辨識和結合到特定的DNA序列,在細菌中同時調控與代謝相關的基因簇表達,從而實作對代謝途徑的整體調節。
此外,鏈黴菌中存在豐富的第2信使系統,如環磷酸腺苷、高度磷酸化的鳥苷酸分子調控(ppGpp調控)等,這些第2信使能夠與轉錄因子相互作用,調整其親和力或構象,從而調節特定基因的轉錄活性。如環二鳥苷酸(c-di-GMP)調節鏈黴菌的生物膠囊形成、生物膜合成和細胞的運動能力,高水平的c-di-GMP可促進生物膠囊的形成和生物膜的合成,從而抑制鏈黴菌的移動和生長,而低水平的c-di-GMP則與鏈黴菌的生長和分化過程相關。C-di-AMP可調節鏈黴菌的細胞壁合成和細胞分裂,其在鏈黴菌的生長和分化階段發揮重要作用。ppGpp的合成與生長階段和菌絲形成密切相關,高水平的ppGpp能夠抑制鏈黴菌的細胞分裂和生長,導致細菌進入休眠狀態[71-73]。
鏈黴菌的轉錄因子調控對於合成具有藥用活性和工業套用前景的代謝產物非常重要,研究鏈黴菌中轉錄因子的結構、功能和調控機制有助於理解鏈黴菌的代謝途徑調控和生物學過程,為最佳化微生物發酵過程和開發新型抗生素等代謝產物提供理論和實踐指導。
綜上,非靶向啟用沈默基因簇的方法是透過調控細胞內的代謝調控因子或環境條件,使沈默的基因簇得以啟用、表達並合成次級代謝產物,涉及一系列復雜的基因調控網絡,調控因子主要包括轉錄因子、訊號分子、蛋白質等。這些調控因子透過與DNA結合或參與訊號傳導途徑,調節鏈黴菌的基因表現水平糊代謝途徑,同時,非靶向調控在細菌生存和對環境的適應過程發揮重要作用,可以幫助細菌響應環境變化或細胞內部訊號,進而調整細菌細胞生長、代謝產物合成和細胞生理狀態。
2.2 靶向調控策略
靶向調控策略是指透過針對特定基因、途徑或調控網絡的操作,實作對生物體的生物學特性進行精準調控以實作特定目的方法。這種策略廣泛用於代謝工程、合成生物學、醫藥科學等領域,能夠改善生物體的效能、代謝途徑和產物合成[74-76],主要透過核糖體工程、啟動子改造、沈默BGCs的異源表達或編輯等方法來實作
2.2.1 核糖體工程
核糖體工程本質是抗生素脅迫目的菌株發生定向突變的一種微生物育種技術,目的是透過改變核糖體的結構和功能,使其能夠最佳化蛋白質合成的速度、準確性和產量[77-78],主要方法為利用一些能夠作用於核糖體和RNA聚合酶等轉錄轉譯元件上的抗生素,促使菌株發生抗藥性突變,達到調控細胞內沈默BGCs表達並促進相關活性次級代謝產物的合成[79]。該方法簡單高效,且可實作對目的基因的精準調控,幾乎可用於所有放線菌菌株中次級代謝產物的挖掘或生物合成過程強化,所選抗生素尤以利福平糊鏈黴素最為廣泛,也是調控機制研究最為透徹的2種抗生素[77]。
利福平誘導放線菌產生的最常見突變位於 rpoB 編碼的RNA聚合酶β-亞基保守的第437位的His/H,該位點容易突變為Tyr/Y、Arg/R和Leu/L等,該突變極易啟用沈默BGCs從而促進新穎次級代謝產物合成,此外rpoB的433位與442位的Ser/S、422位的Leu、424位元的Gln/Q及427位的Asp和440位的Arg也會發生相應突變,而鏈黴素誘導放線菌產生的突位點常見於 rpsL 編碼的核糖體蛋白S12的第88、43位的Lys/K[80-81]。常見的突變形式為Lys88Glu、Lys88Arg和Lys43Asn[82]。Zhang等[83]透過使用利福平300 μg/mL誘導鏈黴菌 S. somaliensis SCSIO ZH66得到了耐藥菌株ZH66-RIF1,相比於野生型菌株在其發酵產物中得到化合物 法雷迪黴素A ,最終確定為RNA聚合酶β亞基Arg444His的突變導致負責雷戴歷黴素A合成的BGCs被啟用。
2.2.2 啟動子改造
具有良好表征的啟動子是代謝工程和合成生物學的重要工具[84]。在重建微生物生物合成代謝途徑改造過程中,常用的方案是利用不同強度的啟動子對目的基因的表達效率進行精細調控,從而提高次級代謝產物的合成效率[85]。常見的鏈黴菌啟動子改造策略包括啟動子替換、引入輔助調控序列和隨機突變、啟動子工程等。
啟動子替換是指透過基因複制和DNA重組技術將目標基因的原有啟動子序列替換為具有不同轉錄調控特性的啟動子序列,替換後的啟動子可以來自於鏈黴菌中其他基因的啟動子或其他物種的啟動子,如促進轉錄的強啟動子,或抑制轉錄的弱啟動子等。向啟動子中引入輔助調控序列是指透過在目標基因啟動子中引入額外的轉錄調控序列,如轉錄因子結合位點、響應元件或強化子序列來增強或抑制目標基因表現的方法,這些輔助調控序列可以來自於鏈黴菌中已知的調控元件,或是從其他物種中獲得。
啟動子隨機突變是指透過隨機引入點突變或插入突變來改變目標基因的啟動子序列,進一步透過篩選或選擇尋找具有不同表達水平的突變體,該方法可以幫助研究者尋找到合適的啟動子序列,從而達到期望的基因表現水平。啟動子工程是指透過基因組編輯技術,如成簇的規律間隔的短回文重復序列系統9(CRISPR/Cas9)系統可直接對鏈黴菌的基因組進行編輯,改變特定基因的啟動子序列,從而實作對特定基因表現的調控,該方法可以精確修改鏈黴菌的啟動子,以達到所需的表達水平[86-88]。
如Li等[89]將天藍色鏈黴菌 S. coelicolor 中的天然啟動子kasOp-受 S. coelicolor 藍色素調節劑(ScbR)和ScbR2調節劑嚴格控制的暫時表達啟動子改造成強啟動子kasOp3,並透過隨機突變和突變文庫篩選消除kasOp3核心啟動子區域內的另一ScbR結合位點獲得了強啟動子kasOp*,並在3種鏈黴菌中比較了kasOp*與另外2種已知強啟動子ermEp*和SF14p的活性,發現kasOp*在3種宿主中均表現出最強的轉錄和轉譯活性。此外,Li等[89]還探索了誘導型啟動子調控BGCs表達的效應,發現與組成型啟動子相比,誘導型啟動子可顯著提高 S. coelicolor 中放線菌紅素和異質土黴素的滴度。上述研究證明啟動子的設計可為鏈黴菌次級代謝產物合成途徑沈默BGCs的表達發揮顯著調控效應。
2.2.3 沈默BGCs的異源表達
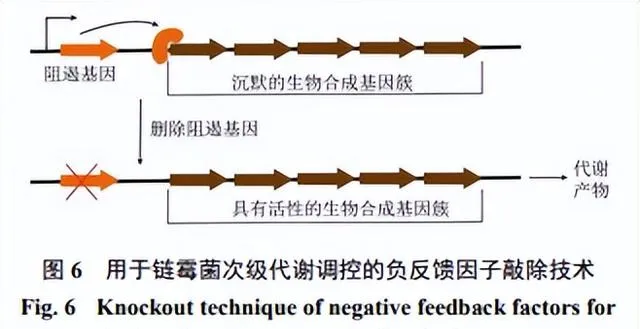
BGCs的「沈默」狀態通常是由於受到直接或間接的阻遏造成,因此去除或「繞過」阻遏因子是啟用沈默BGCs的關鍵。透過生物資訊學工具預測、複制目的基因、修飾基因簇、構建表達載體、轉移基因簇、功能性表達基因、比較分析代謝產物圖譜、最佳化表達等而實作目標BGCs的啟用,這種研究手段稱為BGCs的異源表達[74,90]。選擇合適的宿主是異源表達成敗的關鍵。異源宿主相較於天然宿主具備多種優勢:首先,異源宿主培養條件簡單,其調控體系相對清晰;其次,轉錄元件在異源宿主中更易於控制,使直接複制或重構BGCs後進行異源表達成為可能,透過這種方法,可以規避原宿主中復雜的調控體系(且多為未知),從而啟用沈默BGCs,並產生相應的次級代謝產物[91]。
目前廣泛使用的異源宿主包括大腸桿菌、酵母、鏈黴菌等。大腸桿菌作為常見的細菌宿主,具有高表達能力和培養條件簡單等特點,被廣泛用於重組織蛋白表達[92];而酵母是一種單細胞真核生物,具有較高的蛋白質折疊和修飾能力,常用於復雜蛋白質表達和進行糖基化修飾的研究[93-94]。鏈黴菌具有許多有益特點,如廣泛的物種多樣性、天然的次級代謝能力、高效的蛋白表達系統及豐富的分泌途徑,使其成為許多天然產物BGCs異源表達的理想宿主,如天藍色鏈黴菌、淺紫灰鏈黴菌 S. lavendulae 和白色鏈黴菌 S. albus [95]。Pait等[96]將 S. lavendulae FRI-5的 lav 基因簇在工程菌株 S. avermitilis SUKA22異源表達時發現了一個新的二醇聚酮化合物。
透過異源表達BGCs,可以探索新的產物合成途徑、增加產物的多樣性,並幫助理解和利用BGCs在不同宿主中的表達特征,然而異源表達仍然面臨一些挑戰,如調控異源基因的表達、代謝負擔和代謝通路中間產物的調控等,因此,在設計和實施異源表達策略時,需要仔細考慮特定的研究目的和宿主的適應力。
2.2.4 沈默BGCs的編輯
沈默BGCs的編輯通常是透過敲除或抑制特定基因或基因簇的表達來實作(圖6),常見的基因編輯技術包括CRISPR/Cas9、RNA幹擾(RNAi)、轉基因技術、編輯酶等相關技術[97]。CRISPR/Cas9技術可以精確地敲除或編輯目標基因,利用CRISPR和Cas9核酸酶的復合物,透過編程的方式精確定位目標基因序列,並進行切割、插入或修復等操作,具有操作簡單、高效率和多樣性等優點[98]。
劉昭遠[99]透過CRISPR/Cas9技術敲除海洋放線菌 S. sp. HNS05中的10個基因簇,經對所獲得的突變株代謝產物進行分析發現其中3個基因簇的敲除可抑制該菌株分泌的抑菌活性物質的生物合成。RNAi是一種透過轉錄小幹擾RNA(siRNA)或小發夾RNAs(shRNA)來抑制基因表現的方法。透過設計特異性的siRNA或shRNA序列靶向特定基因或基因簇的mRNA,RNAi技術可以誘導降解目標mRNA,從而沈默基因簇的表達[100]。
轉基因方法透過構建含有負向調控元件(如啟動子或反義序列)的轉基因載體,將其匯入目標生物體內。這些負向調控元件可以抑制目標基因或基因簇的表達,從而實作沈默效果[101]。編輯酶相關技術包括轉錄啟用因子樣效應核酸酶(TALENs)和鋅指核酸酶(ZFNs)。TALENs和ZFNs是一類專門設計的DNA結合蛋白,可以與特定基因序列結合並誘導DNA斷裂,透過設計這些編輯酶靶向基因簇的特定區域,實作基因簇的敲除或編輯[102-103]。
3 結語與展望
鏈黴菌是一類重要的原核微生物,具有豐富的次級代謝產物合成能力,其代謝調控是一個重要而復雜的研究領域。基於鏈黴菌基因組學和轉錄組學的分析表明,鏈黴菌次級代謝產物的生物合成受到一系列基因調控系統的精密控制,這些調控系統涉及到多個層面的調控,包括轉錄調控、訊號傳導、基因組和表觀遺傳學調控等。因此,了解這些調控機制對於挖掘鏈黴菌次級代謝產物生物合成潛力、最佳化產物產量及開發新的活性天然產物均具有重要意義。
在轉錄調控層面,數以百計的轉錄因子被發現與鏈黴菌次級代謝產物的生物合成相關,這其中包括正調控和負調控因子,它們透過結合特定的DNA序列、調節基因轉錄水平、影響酶活性等方式來調節次級代謝途徑關鍵基因的表達。此外,訊號傳導系統在鏈黴菌次級代謝產物生物合成的調控中也發揮重要作用,這些訊號傳導系統包括多種訊號分子的感知、轉導和調控機制。如透過環境刺激、生理訊號、細胞周期等,鏈黴菌可以調節次級代謝途徑的啟動和關閉,從而合理調控產物的合成。
另外,基因組和表觀遺傳學調控在鏈黴菌次級代謝產物生物合成中也被廣泛研究,鏈黴菌具有較大的基因組和豐富的表觀遺傳修飾系統,這些修飾可以在細胞內部、細胞間傳遞並影響細胞內的基因表現和代謝調控,基因組和表觀遺傳學變化可以改變次級代謝途徑中關鍵基因的表達,從而影響次級代謝產物的產生。透過基因工程和代謝工程的手段,也可以對鏈黴菌的代謝通路進行最佳化和改造。這些工作主要涉及調節基因表現水平、增強代謝途徑酶活性、最佳化受質供應等策略。透過這些手段,大量的鏈黴菌菌株次級代謝產物的合成水平被顯著提高。
盡管如此,目前對鏈黴菌代謝調控的研究和套用仍面臨很多挑戰。首先,鏈黴菌生命周期復雜,其代謝調控涉及多個層面,因此,需要深入研究不同階段的鏈黴菌代謝調控網絡,以更好地理解該類別微生物的代謝途徑和調控機制。此外,傳統的發酵調控方法在鏈黴菌代謝工程中的套用也面臨一些限制,這些方法通常耗時且低效,因此需要引入高效、精準的分析、調控技術來加速代謝工程實驗探索的最佳化。數據科學、人工智能和機器學習等方法可以幫助更好地分析和預測代謝調控網絡,從而加速最佳化過程。此外將鏈黴菌代謝調控與合成生物學、代謝工程和先進的生物工程技術相關工具結合,進而嘗試更強大的科研工具和策略開發,將有望為鏈黴菌次級代謝產物的挖掘與生物合成過程強化進一步助力。
綜上,鏈黴菌次級代謝產物的生物合成受到多種調控系統的精密調控,理解這些調控機制有助於深入揭示鏈黴菌次級代謝途徑的調控網絡,進而為最佳化產物產量和開發新的生物活性化合物提供理論指導和實踐基礎。
來 源:孟俊華,張少航, 魯曼霞, 厲 曉, 龔 婕,王 煒, 崔培梧.鏈黴菌次級代謝產物生物合成的調控研究進展 [J]. 中草藥, 2024, 55(14): 4917-4923.











