编译:微科盟-文冉,编辑:微科盟悄咪咪、江舜尧。
微科盟原创微文,欢迎转发转载,本文来源【基因组Genome】公众号。
导读
提高米粒品质是当代水稻育种的首要目标。日本现代水稻育种培育出了两种不同类型的水稻,即食用稻和酿酒稻,它们具有不同的籽粒特征,这表明变异等位基因在育种过程中受到了选择。鉴于及时有效地鉴定过去育种过程中选择的基因对于未来分子育种至关重要, 研究人员进行了全基因组水平的变异鉴定、全基因组关联分析和基于图谱的克隆研究 。结果显示,研究人员 成功鉴定了影响水稻籽粒性状的两个基因: OsMnS 和 OsWOX9D 。 OsMnS 编码一种甘露聚糖合成酶,可增加胚乳中的白芯频率,这是酿酒的理想性状,但会降低籽粒的外观品质。 OsWOX9D 编码一种禾本科特异性含同源框转录因子,可增加籽粒宽度,提高清酒酿造质量。此外,单倍型分析表明,在现代改良过程中它们的缺陷等位基因在东亚而非欧洲被选育出来。此外,研究人员的分析表明,非洲水稻驯化过程中籽粒甘露聚糖含量的降低也可能是由于育种选择导致的 OsMnS 等位基因缺陷造成的。这项研究不仅揭示了水稻籽粒外观品质与营养之间的微妙平衡,而且基于植物"育种辅助基因组学"的概念,为分离复杂性状的因果基因提供了一种新策略。
论文ID
原名: Integrated genome-wide differentiation and association analyses identify causal genes underlying breeding-selected grain quality traits in japonica rice
译名: 全基因组分化和关联分析鉴定粳稻品质性状育种选择的相关基因
期刊: Molecular Plant
IF: 27.5
发表时间: 2023.9
通讯作者: 孙健,Ko Hirano,Makoto Matsuoka
通讯作者单位: 沈阳农业大学水稻研究所,名古屋大学,福岛大学
DOI: 10.1016/j.molp.2023.09.002
实验设计
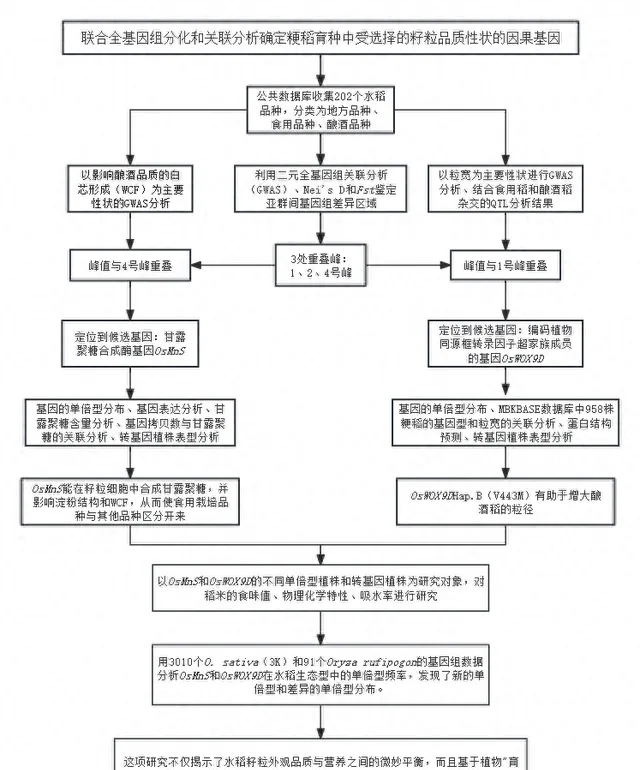
实验结果
1 鉴定区分酿酒水稻和食用水稻的基因组区域
为了鉴定日本水稻育种者所选择的基因,研究人员从公共数据库中收集了202个日本水稻品种,这些品种被归类为现代育种前的地方品种以及用于食用和酿酒的改良品种(图1A)。改良品种是越光(KOS)生产年(1956年)之后培育的品种,当时日本水稻的育种目标从"提高作物产量"转向了"提高籽粒品质"。在该群体中没有观察到明显的亚群体结构。对三个种群对进行了「二元GWAS」(binary GWAS此处命名为"二元GWAS"):"食用品种vs.酿酒品种"、"地方品种vs.食用品种"和"地方品种vs.酿酒品种"。如前所述,通过计算Nei's D和 F st值来估计组间的遗传差异。"食用vs.酿酒品种"的所有三项分析中都观察到了几个共同峰(峰1-4,图1B-1E),并绘制了合理的分位数-分位数(Q-Q)图。 虽然在"地方品种vs.酿酒品种"中没有发现明显的共同峰,但在"食用vs.酿酒品种"中也发现了同在"食用vs.地方品种"中的第1、2和4号峰 (图1E中用红色标出)。这表明 这三个基因组区域是在食用品种而非酿酒品种的育种过程中被特别选择的 。
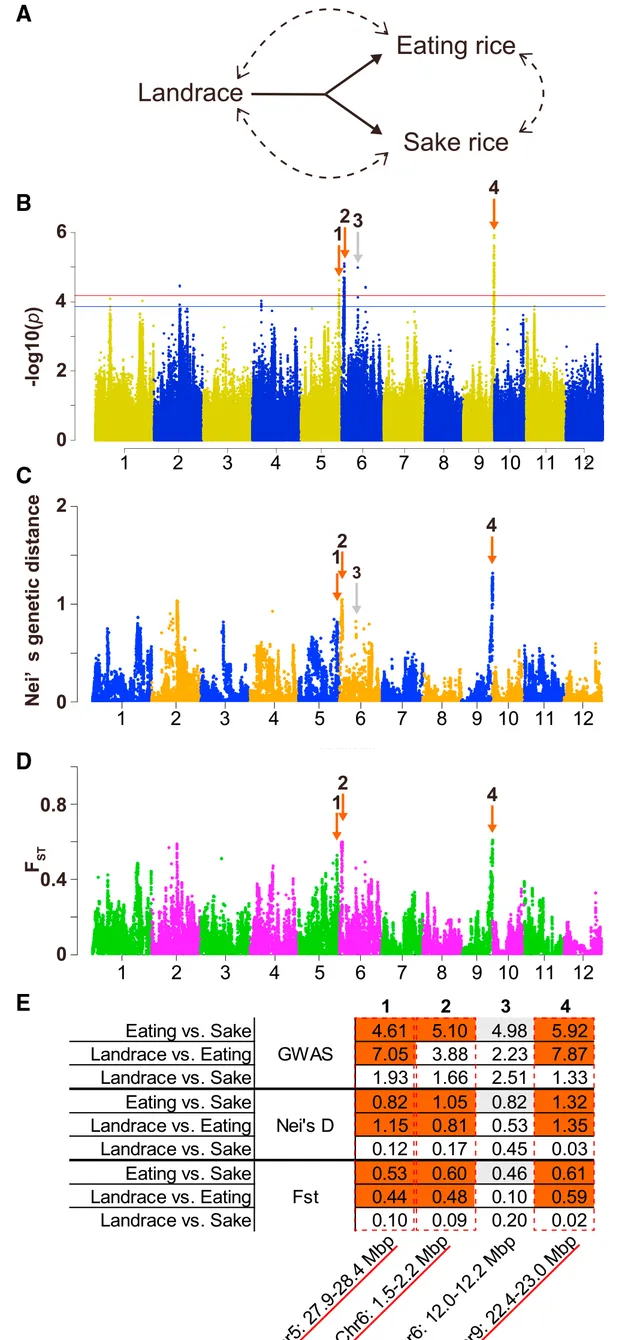
图1| 食用稻和酿酒稻的不同基因组区域。(A)研究人员将日本水稻品种分为三类,即地方品种、食用品种和酿酒品种。(B)二元GWAS的曼哈顿图。(C-D)"食用品种vs.酿酒品种"的全基因组Nei遗传距离(C)和全基因组 Fst (D)。箭头表示在任何分析条件下重叠的先导峰,而红色标出的(1、2和4)表示在三种分析条件下「食用vs.酿酒「和「地方品种vs.食用"的重叠峰。(B)中全基因组显著阈值用水平线表示(红色,0.05/Meff;蓝色,0.1/Meff)。(E)三种分析条件下常见的先导峰列表。列出了每个峰的最高值。红色和灰色方框分别对应于(B-D)中红色和灰色箭头所指的峰值。
2 与食用栽培品种和清酒酿酒栽培品种之间差异相关的基因
米粒中心的白芯形成(WCF)是酿酒大米最重要的性状之一,但对于食用大米来说并不理想。因此,研究人员进 行了一项以WCF为主要性状的GWAS研究 。由于WCF极易受温度影响,研究人员对这一性状进行了多年评估,以尽量减少温度的影响。研究人员根据大比例WCF(LWCF)的百分比对种群进行了分类,并使用132个品种进行了基于LWCF的二元GWAS(图2A)。结果显示出一个主要的峰(图2A中红色箭头所示),该峰与二元GWAS和种群遗传分析发现的4号峰重叠(图1)。因此, 该峰所在处很可能包含一个在酿酒水稻或食用水稻育种过程中筛选出的与WCF相关的基因 。连锁不平衡(LD)分析表明, 候选区域位于第9号染色体(Chr.9)末端的1.1Mb范围内 (图2B和2C)。对候选区域中的所有多态性进行了评估,以确定其对基因所产生的功能影响。在候选区域共发现了6个非同义多态性位点,分别位于6个基因中。通过评估这些突变的功能影响和基因的表达模式,研究人员认为有五个基因不太可能参与WCF,并 最终聚焦于一个基因( LOC_Os09g39920/Os09g0572500 ) ,该基因主要在胚乳和子房中表达,其编码区含有一个10-bp的In/Del(图2D)。该基因编码纤维素合成酶样A(CSLA)的同源物。 日本晴(NPB)参考基因组中的等位基因含有10bp的缺失,不能产生保守的C端区域,而替代等位基因(Hap. B)可以产生该区域。该基因编码的蛋白是β-1,4-甘露聚糖合成酶(MnS)的成员 ,其直系同源物存在于多种植物中。最近,Verhertbruggen等人(2021年)报道了其小麦直系同源物(TaCslA12;TraesCS2B02G560100)在酵母中的表达可导致小麦产生线型甘露聚糖,这有力地表明该蛋白具有甘露聚糖合成酶活性。
日本几乎所有的现代栽培品种都含有这个功能缺失(LOF)的单倍型(Hap.A),而地方品种或酿酒品种大多携带具有生物学功能的单倍型Hap.B(图2E),这表明食用稻选育过程中Hap.A受到了选择。 利用公共数据库检索了 OsMnS 及其直系同源物的器官表达模式,结果显示这些基因主要在发育中的种子中表达,表明该基因能够影响种子特性。由于水稻表达谱数据库是利用携带了Hap.A的NPB构建的,因此,研究人员也检测了该基因在携带Hap.B的酿酒品种Yamadanishiki(YDN)中的表达,结果与NPB一样,YDN的叶片、叶鞘、茎和根中几乎没有该基因的表达。但在开花后7-14天发育中的种子中,YDN中该基因的表达量远高于NPB。 上述基因表达结果表明 OsMnS 在种子发育过程中具有特殊功能 。此外,在水稻叶肉细胞原生质体的内质网中检测到了融合了GFP的OsMnS,而小麦的直系同源物TaCslA12也定位于该细胞。随后,研究人员也测定了种子中的甘露糖含量。与大多数单子叶植物不同,水稻种子中几乎不积累甘露聚糖。但这一观察结果是基于携带Hap.A的水稻品种。事实上, 携带Hap.B的水稻地方品种或酿酒品种中的甘露聚糖含量明显高于携带Hap.A的品种 (图2F)。而其他糖类,如阿拉伯糖、半乳糖、葡萄糖和木糖的含量变化不大。用抗甘露聚糖抗体对稻粒进行原位染色,发现YDN种子的细胞壁上有明显的甘露聚糖信号,在胚乳层上的信号更强,而NPB种子只有微量信号(图2G和2H)。此外,还研究了 OsMnS 单倍型与WCF频率之间的关系。结果显示, 携带Hap.A的品种表现出很低的LWCF值,而在地方品种和酿酒品种中携带Hap.B的品种的LWCF值要高得多 (图2K)。
为了探究 OsMnS 对甘露聚糖合成以及WCF的影响,研究人员将具功能性的 OsMnS Hap.B转入携带Hap.A的NPB植株中。 转基因植株的甘露糖含量更高,植株基因组中 OsMnS 的拷贝数与甘露糖含量显著相关 (图2L;R2=0.8107)。转基因植株的籽粒在原位染色中表现出更高的甘露糖信号(如YDN)和更高的LWCF频率(图2I、2J和2L-2N)。此外,用β-甘露聚糖酶水解米粒发现,转基因植株中的甘露聚糖是同源甘露聚糖,这证实OsMnS可合成甘露聚糖,但不能合成葡甘露聚糖或半乳甘露聚糖。为了评估甘露聚糖对籽粒淀粉的影响,研究人员检测了用OsMnS Hap.B转化的YDN和NPB植株所含的淀粉颗粒。 在这些转基因植株的白芯胚乳中观察到带有气孔的畸形淀粉颗粒,而NPB则表现出有组织的无气孔淀粉颗粒。这与之前的观察结果一致,即白垩化(如白芯)是由畸形的淀粉颗粒造成的 。研究人员还分析了畸形淀粉粒的葡萄糖聚合度,发现与NPB相比,YDN和转基因植株畸形淀粉粒中短链直链淀粉的数量增加,中长直链淀粉的数量减少。此外,为了进一步证实OsMnS的功能,研究人员构建了代表性材料WR04-6的 OsMnS Hap.B突变体。原位染色显示,野生型籽粒中的甘露聚糖信号很强,而突变体籽粒(MNSwr04-6-A和MNSwr04-6-B)中则没有信号。综上所述的研究结果清楚地表明, 4号峰中的 OsMnS 能在籽粒细胞中合成甘露聚糖,并影响淀粉结构和WCF,从而使食用栽培品种与其他品种区分开来。
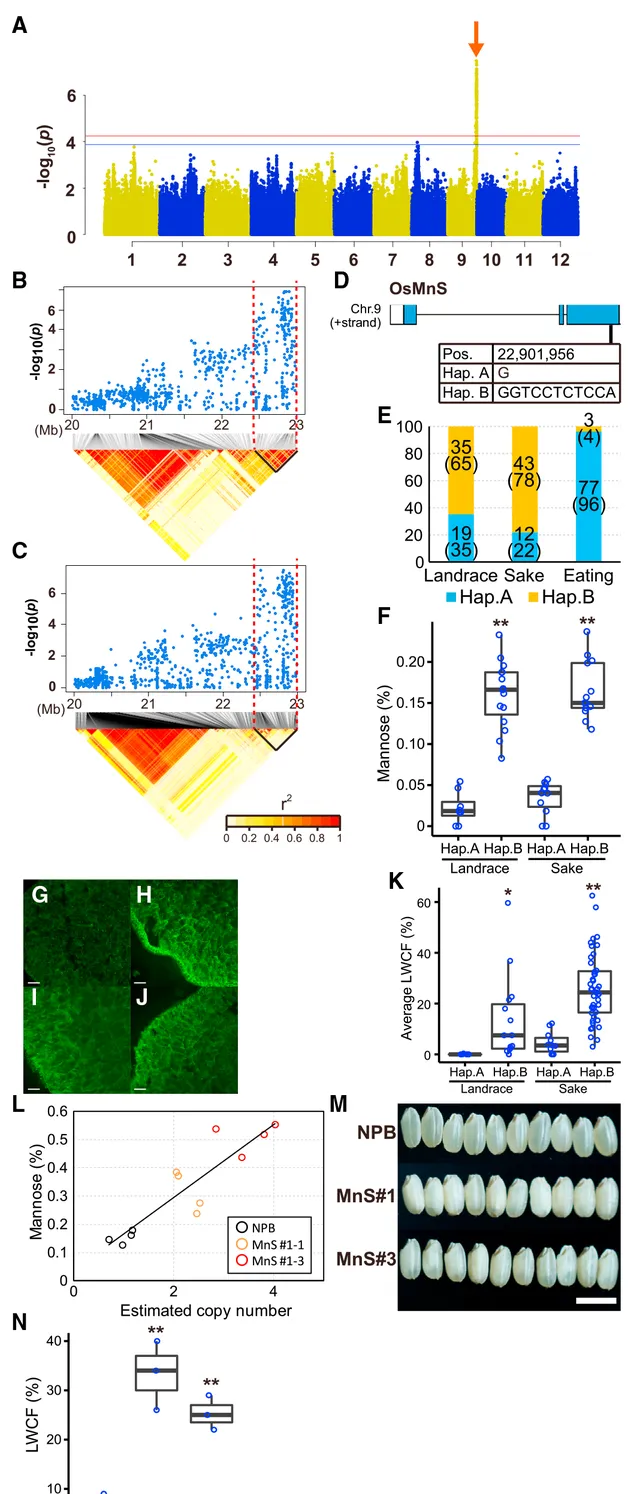
图2| OsMnS 调节WCF和甘露糖含量。(A)LWCF的GWAS。横线表示全基因组显著阈值(红色,0.05/Meff;蓝色,0.1/Meff)。(B-C)围绕图1第4峰(B)和图1A(C)顶峰的局部曼哈顿图和LD热图。(D) OsMnS 的外显子-内含子结构和该基因的DNA多态性。(E) OsMnS 在二元GWAS群体中三个类别的单倍型频率。(F)地方品种和酿酒品种携带Hap.A或Hap.B的水稻之间甘露糖水平的比较( n >8)。通过双尾配对 t 检验,** P <0.01。(G-J)抗甘露聚糖结合蛋白在NPB(G)、YDN(H)、MnS 1号(I)和MnS 3号的稻粒横切面上的原位染色图。实验进行了三次,结果相似。比例尺为50μm。(K)比较携带Hap.A或Hap.B的地方品种和酿酒品种的平均WCF, n >8。通过双尾配对 t 检验,** P <0.01。(L)NPB对照组( n =4)与用功能性 OsMnS Hap.B基因组转化的NPB(MnS 1号的T2世代, n =4)。(M和N)NPB和转化NPB(MnS 1号和3号)的LWCF频率比较。比例尺为5mm(M)。 n =3,NPB中** P <0.01,通过双尾配对 t 检验(N)。
3 控制食用品种和酿酒品种籽粒大小差异的基因
区分食用品种和酿酒品种的另一个性状是籽粒大小,酿酒品种的籽粒比食用品种大。在本研究中, 图1中的1号峰对应于Chr.5的28Mb区域(图3A),它与利用食用稻和酿酒稻杂交进行的粒宽QTL分析中发现的QTL相互重叠 (图3B)。研究人员对粒宽进行了GWAS分析,也在同一区域发现了峰值。接下来,研究人员在KOS背景的水稻(SL-qGWp)中构建了携带YDN qGWp的替换品系,与KOS相比,这些品系的籽粒宽度明显更大(图3C和3D),WCF频率更高,但整体形状没有差异。为了进一步确定该QTL的特征,研究人员对qGWp进行了精细作图,将候选区域缩小到80kb。根据水稻基因组注释项目进行注释(http://rice.uga.edu/),该区域有两个携带SNPs的基因可能会导致氨基酸改变,但其中一个基因(LOC_Os05g48910)被注释为逆转录转座子基因,RAP-DB(https://rapdb.dna.affrc.go.jp/)并未将其预测为一个基因。因此,只有 被注释为 OsWOX9D 的Os05g0564500/LOC_Os05g48990是候选基因 (图3E)。OsWOX9D属于植物同源框转录因子超家族,通过决定细胞命运来调控植物组织和器官的发育。在各种禾本科植物中, OsWOX9D 及其同源物优先在发育中的羽状体和颗粒体中表达,这与决定种子大小的时间和位置非常吻合,支持 OsWOX9D 参与种子发育和种子大小调控。
对来自各种被子植物的 OsWOX9D 同源物的系统进化分析表明, OsWOX9在禾本科植物中分为两个亚群(WOX9D和WOX9A) 。然而,在相同的单子叶植物(如香蕉、扁豆和菖蒲)中并没有观察到这种二分法,这表明在禾本科植物中存在功能分化。对禾本科植物WOX9蛋白的结构进行比较后发现,WOX9D和WOX9A具有共同的残基,也具有只存在于两者之一的残基。食用水稻(Hap.A)和酿酒水稻(Hap.B)之间被取代的氨基酸是V443,它在WOX9D中是保守的,被M取代(图3E)。因此,这一突变预计会影响OsWOX9D的功能。研究人员从MBKBASE数据库(https://www.mbkbase.org/rice)中下载了958株粳稻的基因型和粒宽,并 研究了携带M443的植株与其他携带V443的植株之间的关联 (图3F)。不出所料, 携带M443的植株的粒宽明显大于其他植株 ,这支持了该SNP对粒宽有遗传贡献的假设。研究人员还比较了地方品种与酿酒品种中Hap.A和Hap.B的粒宽(图3G),结果证实Hap.B地方品种的粒宽明显大于Hap.A植株。然而,在一些携带Hap.A的罕见酿酒品种中,其籽粒大小与Hap.B植株相似。这似乎是合理的,因为对于酿酒大米来说,较大的粒径是必不可少的。在这种情况下,更大的粒径很可能是通过其他粒形QTLs的效应实现的,这些QTLs在补图13C所示的GWAS分析中被检测到。最后,研究人员比较了地方品种、酿酒品种和食用水稻的Hap.A频率,如预期的那样, 只有食用稻的Hap.A频率较高(90%),而地方品种和酿酒品种的Hap.A频率低于40% (图3H)。
三维结构预测结果表明,V443M不仅会导致该置换周围的结构发生变化,而且会导致整个蛋白质的结构发生变化。此外,由于之前的研究表明拟南芥WUSCHEL以同源二聚体的形式发挥作用,研究人员利用AlphaFold多聚体预测了OsWOX9D的同源二聚体,发现 V443M位于直接参与二聚体形成的结构域中,可能会引起整个二聚体结构的变化 。酵母双杂交(Y2H)试验证实了OsWOX9D蛋白能够形成二聚体,而且Hap.A二聚体的亲和力明显强于Hap.B二聚体。这表明SNP导致的V443M对OsWOX9D蛋白的生化特性产生了影响。最后,为了直接证实 OsWOX9D 调控籽粒宽度,研究人员构建了两种不同类型的转基因植株。一种是利用CRISPR-Cas9系统在NPB和YDN中破坏OsWOX9D;NPB突变体的粒宽没有变化,而YDN突变体的粒宽有所下降(图3I和3J)。另一种情况发现,用 OsWOX9D Hap.B转化的NPB籽粒宽度有所增加(图3K)。这些结果证实, OsWOX9D Hap.B确实有助于增大酿酒稻的粒径。
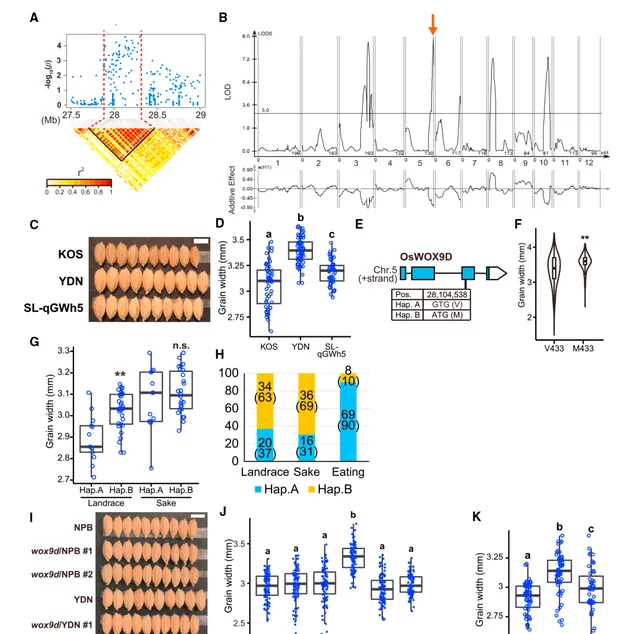
图3| OsWOX9D调节籽粒宽度 。 (A) 图1中峰值1周围的局部曼哈顿图和LD热图。 (B) 用KOS和YDN杂交的RIL对粒宽进行QTL分析。箭头所指为最高QTL,qGWp。 (C和D) KOS、YDN和SL-qGWp之间的粒宽比较。来自YDN的QTL增加了籽粒宽度。比例尺为5mm (C) ; n =100 (D) 。 (E) OsWOX9D 的外显子-内含子结构和该基因的DNA多态性。 (F) 小提琴图显示了带有与Hap.B相对应的V443( n =655)或M443的日本水稻品种的谷物宽度。( n =298)。** P =5.64E-17,通过双尾配对 t 检验。 (G) 方框图显示了携带 OsWOX9D Hap.A或Hap.B的水稻地方品种和酿酒品种的粒宽。 n >13。** P <0.01,通过双尾配对 t 检验。n.s.,无显著差异。 (H) OsWOX9D 在GWAS群体中三个类别的单倍型频率。 (I-J) NPB、YDN和 OsWOX9D 基因敲除植株(wod9d/NPB 1号和2号,以及wox9d/YDN 1号和2号, n =100)的粒宽比较。 (K) 比较NPB、YDN和携带YDN中 OsWOX9D Hap.B基因组区域的NPB的粒宽。 n =40。采用Tukey's HSD检验进行多重比较;不同字母表示在5%水平上的显著性( D 、 H 和 I )。
4 OsMnS和OsWOX9D对食用品质的影响
由于 OsMnS 或 OsWOX9D 的Hap.A在食用稻中被选择,研究人员推测这些基因也与籽粒质量有关。因此,研究人员使用Toyo Taste Mater评估了作为食用稻质量综合指标的食味值,并使用物理测试仪Tensi-pressor评估了米粒的物理化学特性,重点研究了作为熟米饭纹理指标的表面粘性和粘附性。 与携带 OsMnS Hap.A的参考植物NPB相比,Hap.B转化的植物具有明显更高的食味值 (图4A),这可能是由于NPB Hap.B转化植物的熟米饭表面粘性和粘附性较低(图4B-4D)。然而,在具有异质性基因组的本地品种或酿酒水稻品种中没有观察到这种差异。这表明,尽管 OsMnS 对籽粒质量,特别是对籽粒外观有负面影响(图2K-2L),但 OsMnS 对食用品质的影响可能会被一般食用水稻种群中的遗传背景所抵消。
与KOS相比,引入了 OsWOX9D Hap.B的KOS的食味值有所提高(图4E)。然而, OsWOX9D Hap.B的引入没有改变KOS熟米饭的表面粘性或粘附性(图4F-4H),在携带异质性基因组的本地品种或酿酒品种中也没有观察到物理化学特性的差异。另一方面,YND的食味值比KOS差(图4E)。YND产生的米粒比引入 OsWOX9D Hap.B的KOS或KOS要大得多(图3D),这应该有助于改善食味值。尽管如此,YND的食味值较差可归因于其他因素,如表面粘性和粘附性,所有这些因素都远低于KOS(图4F-4H)。
此外,研究人员还直接研究了 OsMnS 和 OsWOX9D 是否影响与清酒生产直接相关的性状。由于吸水量决定了蒸米的消化率,并对发酵有很大影响,因此吸水量是清酒生产中一个非常重要的性状。在浸泡10分钟后测量精米的吸水率。 引入 OsMnS Hap.B能明显提高NPB籽粒的吸水性 (图4I)。用α-淀粉酶处理后蒸米的Brix值(含糖量百分比)能表明米曲霉( Aspergillus oryzae )进行酒精发酵的消化率。 与吸水率结果一致,引入 OsMnS Hap.B提高了NPB籽粒的Brix值 (图4J)。缺乏 OsWOX9D 会降低YDN籽粒的吸水率和Brix值(图4I和4J)。这些结果表明, OsMnS 和 OsWOX9D 的Hap.B能提高发酵速度,使米粒成为清酒的优质原料 。
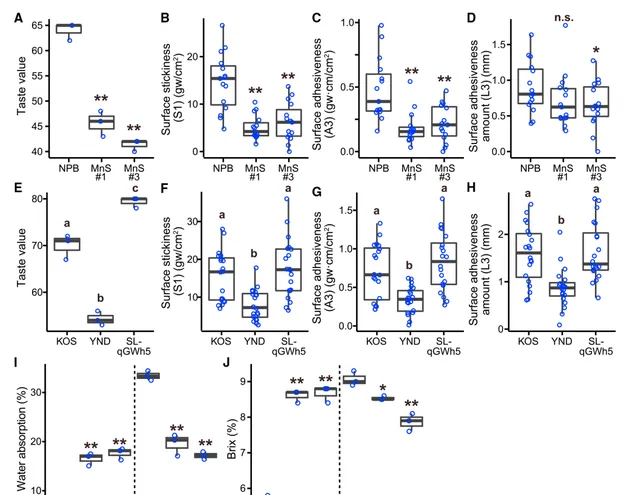
图4| OsMnS 和 OsWOX9D 对籽粒食味质量的影响 。 (A-D) 各图显示了NPB和用 OsMnS Hap.B转化的NPB(MnS 1和3号)籽粒的食味值( A )、表面粘性( B )、表面粘附性( C )和表面粘附量( D )的比较;( A ) n =3,( B-D ) n =15。** P <0.01,* P <0.05;n.s.,通过双尾配对 t 检验无显著差异。 (E-H) 与(A-D)相同,但在NPB、KOS和SL-qGWp之间进行比较。采用Tukey's HSD检验进行多重比较,不同字母表示在5%水平上的显著性。与携带Hap.A的植株相比,通过双尾配对 t 检验后** P <0.01,* P <0.05。 (I-J) 每幅图都显示了NPB和用OsMnS Hap.B转化(左)以及YDN和 OsWOX9D 基因敲除的YDN(右)之间的比较。通过双尾配对 t 检验,** P <0.01,* P <0.05。
5 水稻栽培品种中OsMnS和OsWOX9D的单倍型分布
OsMnS 和 OsWOX9D 的Hap.A单倍型是在日本的粳稻育种过程中被选育出来的。为了研究其他地方或籼稻育种中是否也有类似的选择,研究人员 利用3010个 O. sativa 种质(3K)和91个 Oryza rufipogon 种质的基因组数据分析了它们在水稻生态型中的单倍型频率 。 分析发现在3K组(Hap.C)和 O. rufipogon 组(Hap.D-F)中发现了含有氨基酸交换或插入的新型 OsMnS 单倍型。 研究人员发现,Hap.D-F中的这些突变发生在非保守区域,而Hap.C中的T273K突变位于所有测试的绿色植物(包括绿藻、苔藓和种子植物)中的一个高度保守的残基上。为了证实这些突变的影响,研究人员测量了携带Hap.B-F的植物中的甘露糖含量。 虽然携带Hap.D-F的植株中的甘露糖含量几乎与Hap.B植株相当,但在所有测试的Hap.C植株中仅检测到微量的甘露糖含量(图5A),这表明Hap.C可能是另一种LOF单倍型 。单倍型网络分析表明, Hap.B是原始的单倍型,而Hap.A和Hap.C是由Hap.B突变而来的 (图5B)。扩展单倍型纯合度衰减分析证实了这一点。Hap.A优先出现在粳稻( japonica )中(不包括亚热带地区),Hap.C出现在籼稻( indica )的XI-1中,而这两个单倍型都不在 O. rufipogon 中(图5C)。这表明 粳稻和籼稻XI-1亚群中的Hap.A和Hap.C在驯化后和现代育种过程中分别独立地受到选择 。为了确定Hap.A是否在日本以外的其他地区发生了育种选择,研究人员比较了中国地方品种和现代品种之间的单倍型频率,观察到了类似的趋势(图5D)。事实上,现代品种中的单倍型频率始终高于地方品种中的单倍型频率,中国不同地区的单倍型频率存在一定差异,但由于吉林、辽宁和宁夏不存在地方品种,因此无法进行比较。
研究人员还利用3K种质调查了 Hap.A在粳稻中的全球频率,结果发现了明显的差异 :东亚(EAS)和东南亚岛国(SEAI)的频率大于70%,而南亚大陆(SAC)和欧洲(EUR)国家的频率约为20%,西非国家的频率为60%(图5E)。这可能是由于各地区优势亚群的差异造成的,因此研究人员还比较了亚群之间的Hap.A频率。 温带粳稻主要生长在东亚和欧洲国家,其Hap.A频率有很大差异 :东亚国家为81%,欧洲国家为20%(图5E)。在粳稻混交的地区,虽然品种较少,但也观察到类似的趋势。亚热带粳稻几乎只生长在SAC国家,其Hap.A频率小于10%,而热带粳稻在包括SEAI和西非国家在内的不同地区具有较高的频率(>60%)(图5E)。这些观察结果表明, Hap.A频率取决于亚种群和种植地区,Hap.A受选择过程在东非和东南亚国家积极发生,而在南亚和欧洲国家则没有 。
研究人员还对籼稻中的Hap.C频率进行了同样的分析,由于EUR国家籼稻数量较少,因此忽略了该地区。 EAS国家和菲律宾的Hap.C频率高于其他地区,但这一差异可能是由于XI-1亚群较高所致 ,而XI-1亚群倾向于含有Hap.C。因此,研究人员比较了XI-1亚群中的Hap.C频率,发现EAS国家与其他地区之间没有明显差异(图5F)。这表明,Hap.C频率的差异仅取决于各种植区的XI-1频率,而在XI-1亚群内部没有进一步的选择。事实上,中国的地方品种与现代品种之间的Hap.C频率差异不大(图5G),这表明 与粳稻不同的是, OsMnS 在籼稻的现代育种过程中并不是一个明显的目标。
对于 OsWOX9D ,研究人员发现了新的单倍型——Hap.C-H,在3K种质中有超过10个品种共享该单倍型 (图6A)。与仅在温带粳稻和混交种粳稻中共享的Hap.B不同,Hap.C-E广泛存在于籼稻和粳稻亚群中(图6B),其氨基酸替换位点在禾本科同源物中并不保守。这表明它们不会影响 OsWOX9D 的功能,因此研究人员在随后的分析中将重点放在了Hap.A和B上。3K队列中温带粳稻Hap.B的扩展单倍型纯合度衰减比Hap.A稍慢。这表明Hap.A是一个祖先单倍型,这与Hap.A在所有 O. rufipogon 亚群中都是显性单倍型这一事实相对应(图6B)。相比之下,在日本的栽培品种中,Hap.B的衰减速度比Hap.A快,这与日本最近在水稻育种中选择Hap.A的事实相对应(图3H)。这些观察结果也表明, Hap.B已在温带粳稻中确立并广泛分布,但在现代育种过程中又被祖先Hap.A所取代 。在中国的粳稻中也观察到了同样的现象,几乎所有的粳型杂交水稻都有Hap.A,而>50%的地方品种有Hap.B,现代育成的品种则有>60%的Hap.A(图6C)。在中国的不同地区都观察到了这种重新转换,而在云南和"未知"地区,Hap.A频率有所下降。在EAS(70%)和EUR(30%)国家之间,3K队列中粳稻内的Hap.A/B比率存在显著差异(图6D)。这表明 在现代育种过程中,Hap.B向Hap.A的再转换在EAS和SEAI国家积极发生,而在EUR国家没有发生,这与 OsMnS 的单倍型转换类似 。
最后,研究人员测量了各种野生稻籽粒中的甘露聚糖含量。包括 O. rufipogon 在内的 所有AA基因组水稻的甘露聚糖含量与含有Hap.B的 O. sativa 水稻相似,而其他携带BB、CC、CCDD、EE、FF和HHJJ基因组的野生水稻的甘露聚糖含量要高得多 (>0.4%)(图5A)。研究人员还比较了非洲驯化稻 O. glaberrima 和其祖先 O. barthii 的甘露聚糖水平,两者都携带AA基因组。 O. barthii 的甘露聚糖含量与其他AA型水稻几乎相同,而在一些 O.glaberrima 品种(包括 O. glaberrima 参考品系CG14)中,甘露聚糖含量微乎其微(0.05%),因此这两个物种之间存在显著差异(图5A)。研究人员试图在公开的CG14参考基因组数据中找到 OsMnS 的直系同源物,但未能找到。但成功地将CG14和其他 O. glaberrim a株系的基因组NGS读数映射到IRGSP1.0的 OsMnS 序列,为其同源序列在 O. glaberrima 基因组中的存在提供了强有力的证据。然后,研究人员将CG14的RNA测序读数映射到IRGSP1.0,发现CG14的 OsMnS 因转录错误而失去功能。CG14、 O. barthii 和IRGSP1.0之间的共线性分析进一步证实, O. glaberrima 和 O. barthii 中 OsMnS 的同源序列位于9号染色体的3’端,而该位置在之前的研究中已经报道了存在强烈的驯化信号。这些结果表明, 在非洲水稻驯化过程中, O. glaberrima Chr9的长末端(包括 OsMnS )也可能是驯化的目标 。
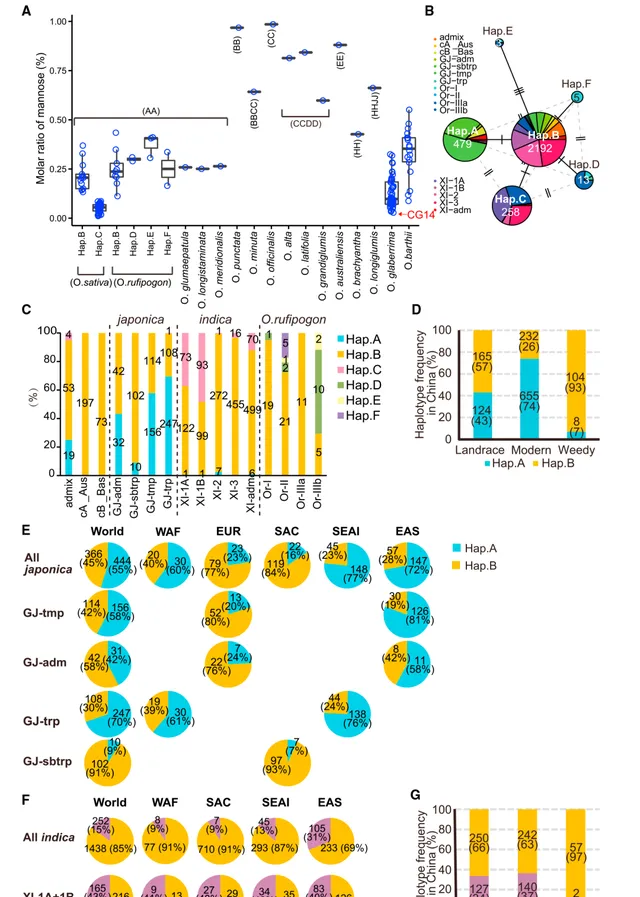
图5| O. sativa 和 O. rufipogon 亚群中 OsMnS 不同单倍型的分布。(A) 在 O.sativa 的Hap.B( n =13)和Hap.C( n =40); O. rufipogon 的Hap.B( n =9),Hap.D( n =2),Hap.E( n =3)和Hap.F( n =2)中的甘露聚糖水平;其他野生水稻种(每个种质 n =1)具有如下的不同基因组:AA( O. glumaepatula 、 O. longistaminata 和 O. meridionalis )、BB( O. punctata )、BBCC( O. minuta )、CC( O. officinalis )、CCDD( O. alta 、 O. latifolia 和 O. grandiglumis )、EE( O. australiensis )、FF( O. brachyantha )和HHJJ( O. longiglumis )。非洲驯化稻 O. glaberrima ( n =48)及其祖先 O. barthii ( n =18)的甘露糖含量。红色箭头表示 O. glaberrima (CG14)的参照物。 (B) OsMnS 的单倍型网络。每个饼中的数字代表加入的数量,亚群的比例用不同颜色表示。 (C) 3K O.sativa 和 O.rufipogon 队列中不同亚群的 OsMnS 单倍型频率。 (D) 中国品种中三个类别的 OsMnS 单倍型频率。 (E) 3K粳稻品种和不同粳稻亚群中 OsMnS 单倍型频率的区域差异。括号中的数值表示品种数量。WAF,西非;EUR,欧洲;SAC,南亚大陆;SEAI,东南亚岛屿;EAS,东亚。 (F) 3K籼稻品种和籼稻品种XI-1(XI-1A+XI-1B)亚群中 OsMnS 单倍型频率的区域差异。
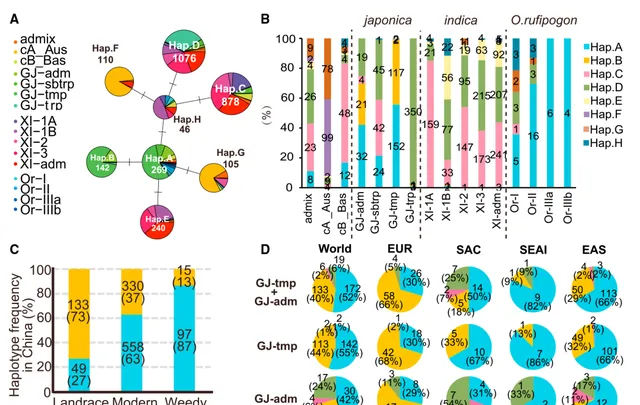
图6| O.sativa 和 O. rufipogon 亚群中 OsWOX9D 不同单倍型的分布情况。(A,B) OsWOX9D 的单倍型网络和单倍型丰度。图示与图5B和5C相同。 (C) OsWOX9D 在中国品种三个类别中的单倍型频率。 (D) 3K粳稻品种和不同粳稻亚群中 OsWOX9D 单倍型丰度的区域差异。括号中的数值表示品种数。
讨论
「育种辅助基因组学":分离因果基因的有效方法
在被不同育种目标选择的群体中,有一些基因和基因位点承受着不同的遗传压力。 本研究利用二元GWAS和群体遗传学方法对日本育种群体(食用稻和酿酒稻)进行了评估,并帮助阐明了 OsMnS 和 OsWOX9D 分别在决定籽粒品质和籽粒大小方面的作用。 尽管对性状的影响仍有待验证,但这种方法比GWAS或QTL分析能更快地识别基因,因此非常有用。此外,研究人员还能利用13年间通过标准化表型方法获得的表型数据确定WCF的QTL。但由于测量困难,所以观测的群体规模有限。即便如此,二元GWAS和群体遗传方法还是让研究人员找到了WCF的相关基因座。这些成功表明,通过育种选择的基因或基因位点不仅可以被有效鉴定,而且即使是测量费时费力的性状,也有可能通过这种方法鉴定出基因位点。
OsMnS 和 OsWOX9D 是籽粒品质育种的靶点
OsMnS 编码甘露聚糖合成酶,是CSLA成员之一。甘露聚糖主要存在于细胞壁中。虽然甘露聚糖是细胞壁的结构成分,也是籽粒中的能量储备,但其生物学功能尚未得到准确表征。在本研究中,研究人员发现 OsMnS 参与水稻WCF、籽粒外观和质地的形成,而这些是决定籽粒品质的重要因素 。淀粉是决定水稻食用品质的关键因素,因为它通过改变淀粉结构直接影响籽粒质地。本研究表明, OsMnS 会影响淀粉的晶体结构和直链淀粉的链长,从而影响WCF和籽粒外观 。细胞壁成分(这里指甘露聚糖)水平的变化不仅决定了籽粒的白垩感,而且还通过改变淀粉结构来影响籽粒的质地和口感。尽管这种影响的分子机制仍然难以捉摸,但研究人员的研究结果表明,细胞壁是决定白垩度、质地和口感的重要因素。另一方面,经常摄入高纤维的饮食有助于维持健康和降低罹患多种疾病的风险。甘露聚糖是一种优质膳食纤维,但随着现代基因改良,大米中的甘露聚糖逐渐消失。不过,它仍然存在于酿酒品种、地方品种和杂交水稻中,这些遗传资源需要得到重视和利用。
研究人员还发现, OsWOX9D 有助于增加酿酒稻的粒宽。在水稻中已经发现了三个WOX9s( OsWOX9D/DWL2 、 OsWOX9A/DWT1 和 DWL1 ),它们相互配合,共同调控植物的生长。之前的研究报道中发现, dwt1/dwl2 双突变体表现出异常的茎秆伸长和圆锥花序发育。而在本研究中, OsWOX9D 单突变体与双突变体观察到的表型并不重叠,这与之前的报道一致。相反, OsWOX9D Hap.B与种子宽度和WCF频率增加有关,这表明 OsWOX9D Hap.B不是一个LOF,而是可能在种子发育过程中独立于 OsWOX9A 起作用。
根据QTL图谱和GWAS, OsWOX9D 是对酿酒稻粒型增加影响最大的QTL (图3B)。然而,用 OsWOX9D Hap.B转化的SL-qGWp和KOS的粒径比YDN小得多(图3D和3I),这表明清酒水稻的粒径变大一定与其他基因有关。为了研究已知的粒度基因是否参与其中,研究人员检测了粒度基因中已知功能核苷酸多态性的存在与否。不出所料,研究人员在酿酒水稻群体中没有发现任何功能核苷酸多态性。由此看来,清酒水稻的大粒型并不完全受已知基因的调控,还受到许多影响微弱的未知基因的影响。
OsMnS 和 OsWOX9D 的进化模型和应用潜力
OsMnS 的Hap.A仅在粳稻育种中被作为目标基因,而其频率在不同种植区有很大差异 (图5E)。这种差异可能是由于各国饮食文化和饮食习惯的不同造成的。例如,欧洲人通常以烩饭、海鲜饭和米饭布丁的形式食用米饭,而东亚人则将米饭煮熟,不加调料用筷子吃饭。此外,SAC国家GJ-sbtrp中Hap.A频率较低(图5E),可能与该亚群中另一个"籽粒品质"基因Waxy的无效等位基因频率异常高(>55%)有关,因为 waxynull 可能会掩盖Hap.A对籽粒品质的影响。事实上,携带功能性 Waxy 的植株的Hap.A频率(14.6%)是 waxynull 植株(5.2%)的三倍。中国不同产区的Hap.A频率也存在差异,除"未知"外的其他产区的Hap.A频率都大于60%(产区不明的品种,即使是现代品种,也可能是很久以前培育的)。这种差异可能是由于Hap.B没有被Hap.A充分取代,而不是由于饮食文化的差异。因此, 用Hap.A替代Hap.B可能是提高Hap.B品种籽粒品质的必要条件 。 Hap.C可能是 OsMnS 的另一个LOF等位基因,只在籼稻的XI-1亚群中发现,这表明它是在XI-1建立期间或之后被选育出来的 。然而,该单倍型不太可能是在育种过程中被积极选育的,因为其单倍型频率在中国籼稻陆生品系和现代品种之间并无显著差异(图5G)。然而,Hap.C品种的主要种植区是EAS国家和菲律宾,这与Hap.A品种的主要种植区重叠,这表明这些国家的人们可能更喜欢用LOF OsMnS 单倍型来培育籼稻和粳稻品种。在这种情况下,Hap.C可能有助于提高籼稻籽粒的品质。
这项研究强调了水稻籽粒中甘露聚糖含量的下降不仅对 O. sativa 而且对 O. glaberrim a的驯化和多样化都很重要。本研究结果表明, 含有AA基因组的野生稻的甘露聚糖含量远远低于含有其他基因组的野生稻 (图5A),这表明水稻属的进化涉及到籽粒中甘露聚糖含量的巨大变化,这可能会导致种子成熟和萌发过程中的生理差异。水稻籽粒中的甘露聚糖在植物生理方面的生物学意义仍有待研究。
OsWOX9D 是本研究中发现的另一个调节籽粒宽度的基因 。与 OsMnS 一样,它的单倍型转换也显示出Hap.B被Hap.A取代(图3F和6C),而且Hap.A的频率远高于欧洲国家(图6D)。虽然这一趋势不如 OsMnS 那么明显,但有可能是 OsWOX9D 的Hap.A与 OsMnS 一样, OsWOX9D 的Hap.A在EAS国家被优先选择 ,可能是为了减少WCF。相反,由于Hap.A是祖先类型,而Hap.B只在温带地区发现,因此Hap.B只在温带粳稻中建立和传播。虽然在驯化过程中选择的衍生单倍型在现代育种中恢复到其祖先类型的情况非常罕见,但当现代育种的目的与驯化过程的目的不同时,这种现象可能会发生。











