編譯:微科盟 伊一,編輯:微科盟 景行、江舜堯。
微科盟原創微文,歡迎轉發轉載,轉載請註明來源於【轉錄組】公眾號。
導讀
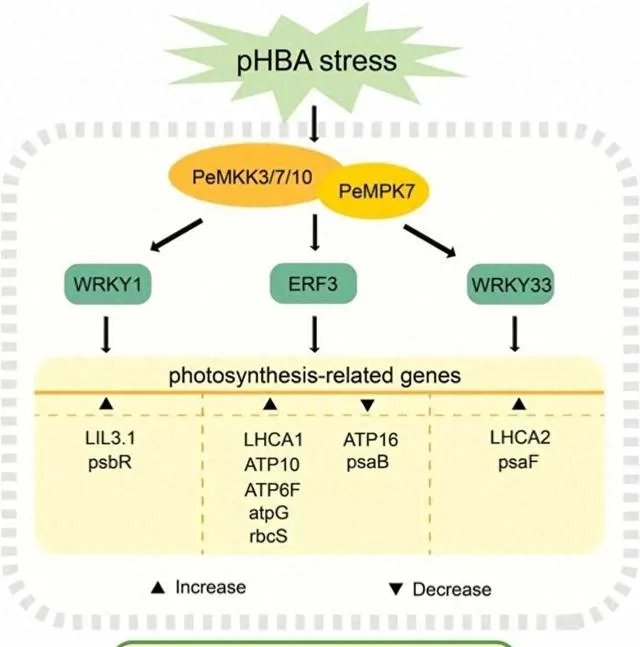
由於楊樹的連續種植,其生長和生物量積累可能會受到土壤中對羥基苯甲酸(pHBA)等等位化學物質積累的負面影響。光合作用是植物最基本的過程,會受到pHBA脅迫的負面影響。因此,提高pHBA脅迫下的光合作用能力對促進楊樹的生長至關重要。絲裂原活化蛋白激酶(MAPK)級聯途徑廣泛參與植物的環境脅迫響應。然而,在pHBA脅迫下,MAPK通路基因對光合作用相關通路的調控機制尚不清楚。 本研究透過轉錄組分析和加權基因共表達網路分析觀察到,PeMPK7在楊樹中過表達可調控pHBA脅迫早期光合作用相關基因和轉錄因子基因(即WRKY1、WRKY33和ERF3)的表達 。此外, PeMPK7還能改善楊樹在長期pHBA脅迫下的光合作用 。此外, 酵母雙雜交和拉下試驗證實了PeMPK7和PeMKK7/10之間的交互作用 。根據這些結果, 本研究構建了PeMPK7參與調控光合作用的途徑示意圖 。本研究為透過MAPK級聯途徑調控pHBA脅迫的分子機制提供了新的見解。
論文ID
原名: Potential molecular mechanism of photosynthesis regulation by PeMPK7 in poplar under para-hydroxybenzoic acid stress
譯名: 對羥基苯甲酸脅迫下PeMPK7調節楊樹光合作用的潛在分子機制
期刊: Ecotoxicology and Environmental Safety
IF: 6.2
發表時間: 2024年4月
通訊作者: 國靜
通訊作者單位: 山東農業大學林學院
DOI號: 10.1016/j.ecoenv.2024.116329
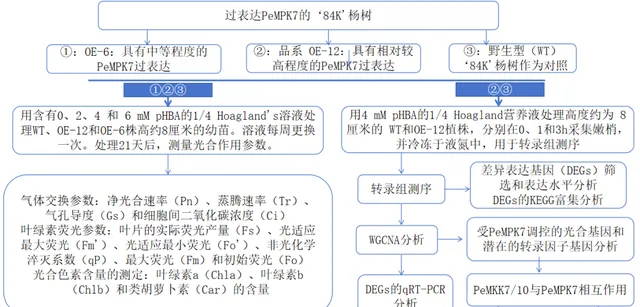
實驗設計
結果
1 過表達PeMPK7調控pHBA脅迫下光合作用相關基因的表達
研究者之前的研究報道,PeMPK7能增強楊樹對pHBA脅迫的耐受性。 為了研究PeMPK7過表達後楊樹早期耐受pHBA脅迫的分子機制,研究者用4mM pHBA處理高度約為8cm的幼苗。1小時後,WT和OE-12植株出現莖尖彎曲和脫水現象 。然而,與WT植物相比,過表達PeMPK7的植物受到的損害要小得多。 處理3小時後,雖然所有植株的莖尖彎曲和葉片脫水程度都有所增加,但WT植株比OE-12植株受到的傷害更大 (補充圖S1)。因此, 研究者在對WT和OE-12幼苗施加pHBA脅迫0、1和3小時後,對其葉片進行了轉錄組測序分析。 這些純凈數據被合並成35422個unigenes,平均長度為1106.93bp,總長度為61984637bp。所有全長轉錄本都在GO、KEGG、Pfam、Swissprot、eggNOG和NR資料庫中進行了註釋。
研究者以 p< 0.05和|log2-foldchange|>1作為篩選差異表達基因(DEGs)的標準。 研究者對篩選的DEGs的表達水平進行了分析 (圖1A)。 在沒有pHBA的情況下,PeMPK7高表達植株(T0)和WT植株(WT0)之間有1659個DEGs(894個上調,765個下調)(T0-vs-WT0) 。 pHBA處理1小時後,PeMPK7基因過表達植株(T1)與WT植株(WT1)之間出現了1671個DEGs(1005個上調,666個下調)(T1-vs-WT1) 。 pHBA處理3小時後,PeMPK7基因高表達植株(T3)與WT植株(WT3)之間出現了1890個DEGs(上調1069個,下調829個)(T3-vsWT3) 。總體而言, 隨著pHBA處理時間的延長,PeMPK7高表達植株與WT植株之間的DEGs數量有所增加 。

圖1.pHBA處理0、1和3小時後差異表達基因(DEGs)的韋恩圖。 (A)PeMPK7過表達(T)和野生型(WT)植株的所有DEGs。(B)T和WT植物光合作用相關DEGs。
研究者在T0-vs-WT0、T1-vs-WT1和T3-vs-WT3對比組中,分別發現了73、52和50個與光合作用相關的DEGs (圖1B)。如補充表S2所示, 各對比組分別有66、25和32個基因上調,這些基因主要與采光復合體I/II葉綠素a/結合蛋白、光系統I/II亞基和磷酸布洛激酶有關 。 各對比組分別有7、27和18個基因下調,主要與V型H+轉運ATPase亞基C、磷酸甘油酸激酶和光系統I組裝蛋白有關 。總體而言, 與WT植株相比,70.9%的光合作用相關基因在過表達PeMPK7的楊樹葉片中顯著上調 ( p< 0.05)(圖2),這些基因主要與光系統I和II亞基以及光合葉綠素結合蛋白有關。因此, 研究者推測過表達PeMPK7的楊樹植株比WT植株捕獲光能的能力更強。
圖2. 在pHBA處理0、1和3小時後,PeMPK7-高表達(T)植株與野生型(WT)植株之間光合作用相關差異表達基因的熱圖(A)。(B)與光合系統I和II有關的基因。(C)參與卡爾文迴圈的關鍵酶。
此外, 研究者還利用KEGG富集分析研究了DEGs參與的主要光合作用途徑 。 與光合作用相關的DEGs的兩個最顯著富集途徑是"光合作用(ko00195)"和"光合作用觸角蛋白(ko00196) "。此外,在"光合生物的碳固定(ko00710)"等通路中也富集了DEGs(圖3B)。
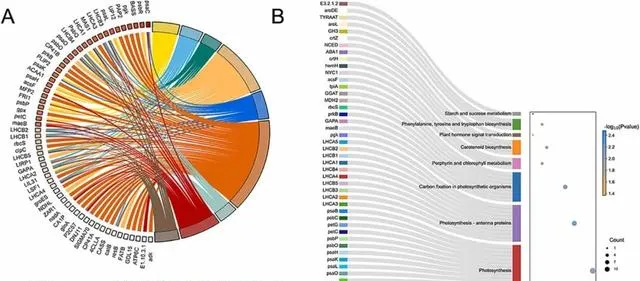
圖3.與光合作用相關的差異表達基因(DEGs)的GO和KEGG分析。 (A)共有9個GO術語與DEGs相關。(B)共有8個KEGG通路與DEGs相關。氣泡圖中圓點的顏色和大小分別代表P值和基因數。
2 WGCNA揭示了光合作用的樞紐基因
研究者根據轉錄組中的基因,進行了WGCNA分析(圖4),共產生了9個共表達模組,每個模組包含93-5880個基因 。在這9個模組中,有5個模組包含超過1000個DEGs,分別是綠松石模組(5880個DEGs)、藍色模組(2417個DEGs)、棕色模組(2093個DEGs)、灰色模組(3495個DEGs)和黃色模組(1313個DEGs)。 在這些模組中,有三個模組與光合作用的相關性最高,即綠松石模組、棕色模組和藍色模組,分別有30個、21個和14個光合作用相關基因。因此,研究者選這三個模組作進一步分析的物件。
研究者透過構建共表達網路,確定了每個模組中的樞紐基因 。 綠松石色、棕色和藍色模組分別包括5個(LHCA1、PetA、psaB、psaG和psbK)、4個(ATP6C、psbR、FaQR和LIL3.1)和4個(LHCA2、psbW、psaF和CAAT6)中心基因 (圖5C-E)。在上述樞紐基因中, 2個(LHCA1和LHCA2)富集於光合天線蛋白通路,6個(psaB、psaG、psbK、psbR、psbW和psaF)富集於光合作用蛋白通路。研究者推測,這8個與光合作用相關的基因可能有助於在PeMPK7過表達楊樹應對pHBA脅迫時維持高水平的光合作用。
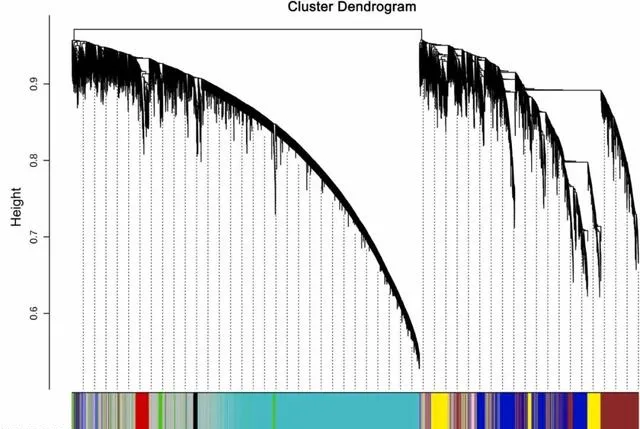
圖4.顯示加權基因共表達網路辨識的共表達模組的層次聚類樹。
3 受PeMPK7調控的光合基因和潛在的轉錄因子基因
研究者根據三個模組的共表達網路,透過分析網路節點關系,構建了光合作用相關子網路 (圖5A)。該子網路主要由光合作用相關基因組成,這些基因均為非轉錄因子(TFs)及其潛在調控TFs。 透過對子網路中的TF進行基因家族分類,研究者發現共表達網路中的光合作用相關基因主要受WRKY(WRKY1和WRKY33)、ZIP(ZIP7和ZTP2)、ERF(ERF3)、bHLH(PIF4)和bZIP(HY5)基因家族的調控 。隨著pHBA濃度的增加,HY5、ZIP7、PIF4和ZIP2在PeMPK7過表達植株和WT植株中的表達水平都穩步下降。相反,隨著pHBA濃度的增加,WRKY1、WRKY33和ERF3在PeMPK7過表達植株和WT植株中的表達量都有所增加。此外,在pHBA脅迫下,PeMPK7-高表達植株中WRKY1、WRKY33和ERF3的表達水平顯著高於WT植株(圖5B)。 研究者推測PeMPK7可能透過調節WRKY1、WRKY33和ERF3等TFs的表達來積極調節楊樹在pHBA脅迫下的光合作用。
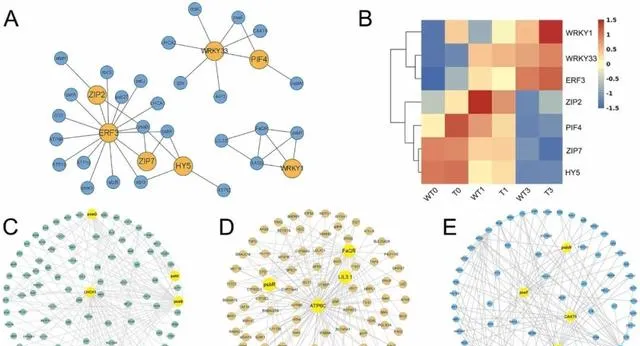
圖5.差異表達基因的共表達網路分析。 (A)與光合作用相關的子網路。橙色圓圈代表轉錄因子(TFs),藍色圓圈代表與光合作用相關的基因。(B)熱圖顯示基因改造植物和WT植物在三個時間點的TFs表達水平。(C-E)黃色圓圈代表樞紐基因。
4 DEGs的qRT-PCR分析
為了驗證轉錄組的結果,研究者從轉錄組數據中隨機選擇了5個與光合作用相關的DEGs(LHCB1、LHCB5、PsaC、PsbR和LIL3.1)和6個其他DEGs(GALE、PP2C、SPG20、PIF4、rpiA和AUX1)進行qRT-PCR分析 。在pHBA處理後0、1和3小時,PeMPK7過表達植株中LHCB1、LHCB5、PsaC、PsbR、LIL3.1和AUX1的表達水平均高於WT植株。在pHBA處理3h後,PeMPK7基因表現植株中GALE、PIF4、PP2C和SPG20的表達水平高於WT植株,而rpiA的表達水平低於WT植株(圖6)。 這11個基因的相對表達水平與轉錄組結果一致,證實了轉錄組數據的準確性 。此外,這些結果表明過表達PeMPK7可提高光合作用相關基因如LHCB1、LHCB5、PsaC、PsbR和LIL3.1的表達水平,使植物在pHBA脅迫下保持較高的光合速率。
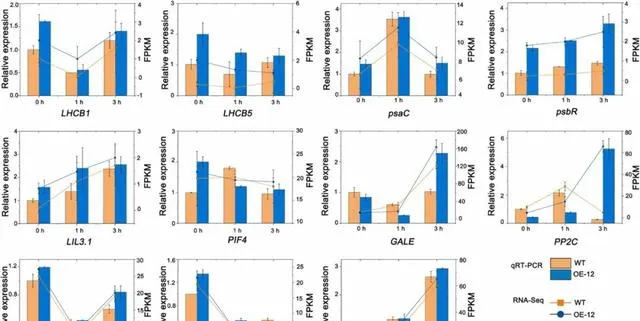
圖6.轉錄組中差異表達基因的qRT-PCR驗證。數據以三個重復的平均值±SD表示。
5 過表達PeMPK7可提高pHBA脅迫下楊樹的光合作用
為了進一步研究PeMPK7是否參與調節楊樹在長期pHBA脅迫下的光合作用,研究者對OE-6、OE-12和WT植物分別用0、2、4和6mM pHBA處理。在pHBA處理的第21天,測量植株的光合特性。
pHBA處理前,OE-6和OE-12中Chla、Chlb和Car的含量顯著高於WT植株,但OE-6中Chlb的含量除外( p< 0.05)。 經pHBA處理後,三種植物中Chla、Chlb和Car的含量均以濃度依賴性的方式下降 。 經6mM pHBA處理後,三種植物中Chla、Chlb和Car的含量最低 。然而, OE-12和OE-6植株中的含量高於WT植株 (圖7)。與暴露於0mM pHBA的植株相比,OE-6、OE-12和WT植株的Chla含量分別降低了21.15%、27.68%和38.83%。因此, 研究者認為pHBA處理抑制了楊樹葉片中葉綠素的生物合成或加速了葉綠素的降解 。然而, 在楊樹中過表達PeMPK7可促進Chla、Chlb和Car的合成或緩解它們的降解。
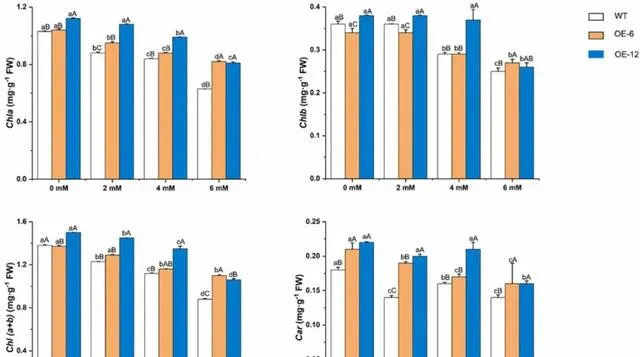
圖7.pHBA脅迫對過表達PeMPK7植物葉片光合色素含量的影響。 數據以三個重復的平均值±SD表示。不同字母表示差異顯著( p< 0.05)。
在pHBA脅迫處理前,OE-6和OE-12植株的Pn顯著高於WT植株( p< 0.05);但在Tr、Gs、Ci和Ls方面,OE-6、OE-12和WT植株沒有觀察到顯著差異。 經pHBA處理後,隨著pBHA濃度的增加,三種植株的Pn、Tr、Gs和Ls均下降,Ci上升。 經2mM pHBA處理後,OE-12和OE-6植株的Gs和Ls顯著高於WT植株( p< 0.05)。用4mM pHBA處理後,OE-12和OE-6植株的Ls明顯高於WT植株。經6mM pHBA處理後,OE-12和OE-6植株的Pn、Tr、Gs和Ls顯著高於WT植株( p< 0.05),Ci顯著低於WT植株(圖8)。 這些結果表明,過表達PeMPK7能改善高濃度pHBA條件下楊樹的Pn和蒸散作用,並且過表達PeMPK7的楊樹葉片的氣孔和Ci的調節能力優於WT植株。
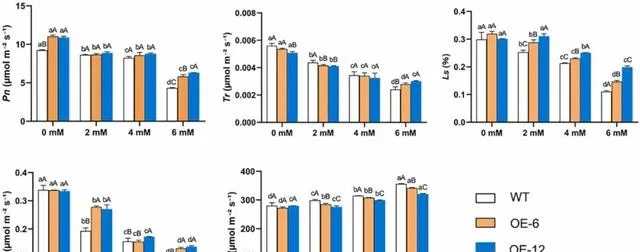
圖8.pHBA脅迫對過表達PeMPK7植物葉片瓦斯交換參數的影響。 數據以三個重復的平均值±SD表示。不同字母表示差異顯著( p< 0.05)。
pHBA處理前,OE-6、OE-12和WT植株的Fv/Fm、Fv/Fo、ΦPSII、qP和NPQ無明顯差異。pHBA處理後,當pHBA濃度低於6mM時,Fv/Fm、Fv/Fo、ΦPSII和qP的值隨pHBA濃度的增加而降低,而NPQ則隨pHBA濃度的增加而升高 。經6mM pHBA處理後,三種植株的Fv/Fm、Fv/Fo、ΦPSII和qP值均最小,OE-12和OE-6植株的Fv/Fm、Fv/Fo、ΦPSII、qP和NPQ值均高於WT植株(圖9)。 這些結果表明,pHBA脅迫顯著抑制了楊樹葉片的光合效率、PSII電位活性、ΦPSII和電子傳遞。 隨著 pHBA濃度的增加,OE-12和OE-6植株的光化學效率、電子傳遞速率、PSII反應中心開放度和調節能力均明顯高於WT植株。
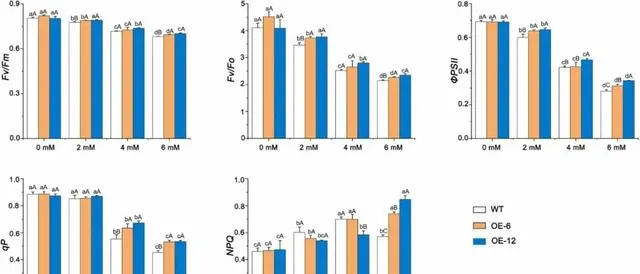
圖9.pHBA脅迫對過表達PeMPK7植物葉片葉綠素熒光參數的影響。 數據以三個重復的平均值±SD表示。不同字母表示差異顯著( p <0.05)。
6 PeMKK7/10與PeMPK7交互作用
MAPK在被其上遊MPKK磷酸化後參與植物防禦反應。研究者之前的研究報道了PeMPK7與PeMKK3之間的交互作用。 為了進一步了解PeMPK7在調控楊樹對pHBA脅迫響應中作用的分子機制,研究者對PeMPK7與其他MPKKs的交互作用進行了研究。 根據STRING資料庫的預測,PeMKK3、PeMKK7、PeMKK9和PeMKK10的得分高於其他MPKKs。因此, 研究者進行了酵母雙雜交和拉下試驗來分析PeMPK7和PeMKK7/9/10之間的蛋白質交互作用。
在酵母雙雜交試驗中,PeMPK7與pGBKT7載體中的GAL4DNA結合域融合,而PeMKK7、PeMKK9和PeMKK10則複制到pGADT7載體中的GAL4啟用域 。結果表明, 陽性對照、陰性對照和pGBKT7-PeMPK7+pGADT7-PeMKK7/9/10共轉化的酵母菌株在SD/-Trp/-Leu固體培養基上生長正常 。除陰性對照外,所有菌株均在SD/-Trp/-Leu/-Ade/-His/X-α-Gal/100ng/mLAbA固體培養基上生長,並在X-α-Gal染色後呈現著色(圖10A)。 這些結果表明,PeMPK7可能與PeMKK7、PeMKK9和PeMKK10交互作用 。 為了進一步證實這一點,研究者使用融合蛋白PeMPK7-His和PeMKK7/9/10-GST進行了拉下試驗 。結果表明 ,在體外,PeMPK7-His與PeMKK7-GST和PeMKK10-GST形成了異二聚體,這表明PeMPK7與PeMKK7/10交互作用,而不是與PeMKK9交互作用 (圖10B-D)。
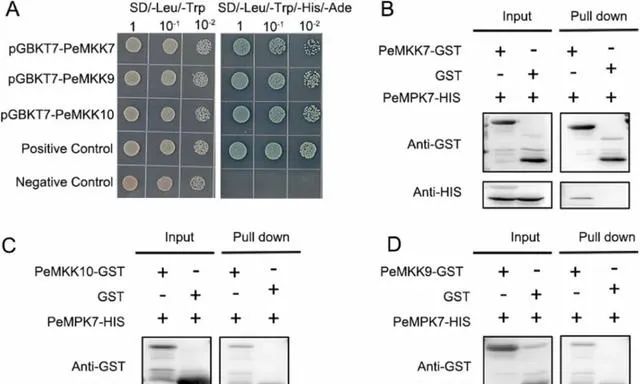
圖10.PeMPK7與PeMKK7/10交互作用。 (A)酵母雙雜交試驗。菌株在SD/-Trp/-Leu和SD/-Trp/-Leu/-Ade/-His/X-α-Gal/AbA平板上生長的能力表示各自的蛋白-蛋白交互作用;1、10-1和10-2代表酵母稀釋度。pGBKT7-53+pGADT7-T和pGBKT7-Lam+pGADT7-T的酵母菌株分別作為陽性對照和陰性對照。(B-D)為驗證PeMPK7和PeMKK7/9/10之間的蛋白交互作用,進行了牽引試驗。在大腸桿菌中誘導PeMPK7-GST和PeMPKKs蛋白,空GST蛋白作為對照。
7 PeMKK7/10的表達受pHBA調控
為了研究PeMKK7/10的表達是否受pHBA的調控,研究者對經pHBA處理的WT植株進行了qRT-PCR。經pHBA處理1、3和6h後,PeMKK7和PeMKK10的表達均被顯著誘導。
討論
光合作用是植物交換瓦斯和合成能量的主要方式。環境因素會影響植物的光合特性。pHBA脅迫會破壞葉片中葉綠體的結構和功能,抑制光合作用,導致植物無法維持正常的生理活動。 本研究透過轉錄組和光合生理指標分析證明,PeMPK7過表達可提高楊樹應對pHBA脅迫的光合作用。
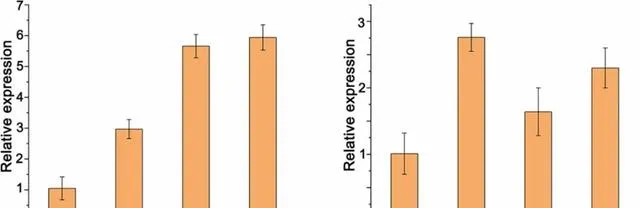
圖11.pHBA處理下楊樹中PeMKK7和PeMKK10的表達分析。 數據以三個重復的平均值±SD表示。
1 PeMPK7參與了pHBA脅迫下楊樹葉綠素和類胡蘿蔔素的合成和代謝
葉片是植物的主要光合器官,對作物的品質和產量至關重要。葉綠素濃度以及葉綠體的形狀和結構直接影響光合作用的效率。葉綠素、碳水化合物、脂質和蛋白質復合物構成了光合系統,這是一個吸收光的功能單元。透過吸收和傳遞能量,葉綠素在光合系統中發揮著特殊而關鍵的功能。此外,葉綠素的生物合成、積累和降解還與葉綠體發育、光形態形成和葉綠體-細胞核訊號傳遞有關。PSI和PSII中的主要色素是Chla和Chlb,它們在Lhc蛋白中捕獲光能。Car在光保護中起著至關重要的作用。在外源FA處理下,草莓葉片中Chla、Chlb和Car的含量下降,導致光合作用速率降低。之前的研究報道,SlMAPK3的過表達增加了番茄的葉綠素含量和光合作用活性。與這些研究結果一致, 本研究表明,在pHBA脅迫下,楊樹葉片中Chla、Chlb和Car的含量顯著下降 (圖7)。此外, 在高濃度pHBA脅迫下,過表達PeMPK7會促進楊樹葉片中Chla、Chlb和Car的積累。同樣,PeMPK7的過表達也影響了參與血紅素和葉綠素代謝以及Car生物合成、運輸和降解過程的基因的轉錄水平 。 在本研究中,與血紅素和葉綠素代謝相關的DEGs在過表達PeMPK7的植株中顯著上調 ,這與Ma等人的研究一致。 在本研究中,研究者透過WGCNA和共表達網路構建分析確定(圖5D)采光復合物樣蛋白3同型1(LIL3.1)為棕色模組中的一個樞紐基因。 采光復合物蛋白可以正向調節葉綠素的生物合成,在光合作用中發揮關鍵作用。 qRT-PCR研究結果表明,在pHBA處理0、1和3小時後,PeMPK7過表達植株的LIL3.1轉錄水平高於WT植株 (圖6)。因此,在pHBA脅迫下,PeMPK7可透過調節葉綠素的合成和代謝以及碳水化合物的生物合成和降解過程來維持基因改造植物的正常光合作用。
2 PeMPK7參與了pHBA脅迫下楊樹的光合碳迴圈過程
光合效率下降是光抑制的最顯著特征,它反映了光合系統的執行狀況。酚類化合物可透過影響膜電位、滲透性和活性來影響細胞膜功能。它們將訊號從根部傳遞到地上部份,影響植物體內的酶活性和滲透脅迫,並導致氣孔關閉,從而導致植物光合速率下降。pHBA能顯著抑制楊樹的光合作用,且抑制作用隨pHBA濃度的增加而增強,其主要原因是非氣孔限制。本 研究結果表明,隨著pHBA濃度的增加,楊樹葉片的Ci增加;Ls顯著降低,Pn逐漸降低 (圖8)。這與之前關於酚酸脅迫下楊樹Pn和Ls下降的研究結果一致。 這表明非氣孔限制是促進Pn下降的主要因素;這可能是由於在pHBA脅迫下葉綠體和類葉綠體結構被破壞,導致光合活動紊亂和葉綠素含量下降 。然而, OE-6和OE-12植株Pn的下降振幅明顯低於WT植株,說明PeMPK7的過表達能有效提高植株的凈光合效率 。此外, 隨著pHBA濃度的增加,楊樹的Tr和Gs逐漸降低。這表明,pHBA脅迫會導致楊樹氣孔關閉、蒸散作用減弱、水分代謝能力下降 。 在高濃度pHBA脅迫下,PeMPK7表達植株Tr的降低程度明顯低於WT植株,說明在高濃度pHBA脅迫下,PeMPK7表達植株的水分代謝能力優於WT植株 。之前有研究報道,在楊樹中過表達PeNAC122能在滲透脅迫下維持較高的瓦斯交換速率。 轉錄組分析表明,在pHBA脅迫下,PeMAPK7過表達植株中NAC家族基因如NAC104、NAC55、NAC2和NAC72的表達水平顯著高於WT植株。 因此, 研究者推測PeMPK7透過調節這些NAC家族DEGs來維持較高的瓦斯交換率 。這一假設需要透過分子生物學實驗進一步驗證。基因改造水稻中Rubisco含量的增加和Rubisco活性的維持可在最適溫度範圍內的中等高溫條件下提高光合作用。同樣, 轉錄組分析表明,與WT植株相比,PeMPK7的過表達顯著上調了植物卡爾文迴圈中編碼酶的大多數關鍵基因,如RbcS、maeB、cynT、can、pgk、prkB和GAPA (圖2C和12)。 研究者推測PeMPK7的過表達可能會顯著提高Rubisco的羧化和RuBP的再生,從而提高植物的光合能力 。結合之前關於PeMPK7改善滲透調節能力的研究, 研究者推測PeMPK7可能會增加滲透調節物質的含量,降低滲透勢,維持細胞的張力壓力,促進植物體內各種物質代謝的順利進行,從而維持較高的光合效率和蒸散速率 ,這有助於植物抵抗pHBA脅迫。
3 PeMPK7參與了pHBA脅迫下楊樹光能的吸收和轉化
葉綠素熒光參數作為植物適應各種環境的內在指標,可用於確定環境脅迫下光合器官的受損程度。例如,Fv/Fm是PSII的最大光化學效率,反映了反應中心的初級光能轉換效率;qP反映了PSII觸角色素吸收的光能中用於光化學電子傳遞的份額;ФPSII反映了PSII反應中心的實際光能捕獲效率。在pHBA脅迫下, Populus × euramericana ‘Neva’和Rumex acetosa的Fv/Fm和ФPSII都有所下降。同樣, 在本研究中,pHBA脅迫後楊樹的Fv/Fm、ФPSII和qP顯著降低。這些結果表明,pHBA脅迫會對光合器官造成損傷。
有趣的是, PeMPK7過表達植株的Fv/Fm遠大於WT植株 (圖9), 且在4mM和6mM pHBA處理後,OE-6和OE-12植株ФPSII和qP的下降振幅遠小於WT植株 (圖9)。 這表明過表達PeMPK7可增加光化學轉移所需的光能,從而提高pHBA脅迫下植物光合電子傳遞的效率 。與研究者的結果一致,在幹旱脅迫下,過表達SlMAPK3的番茄植株的ФPSII和qP值均高於WT植株。由於SlMAPK3和PeMPK7分別屬於A組和C組MAPK, 研究者推測A組和C組MAPK基因在非生物脅迫下具有提高光合電子傳遞效率的保守功能。
在鋅脅迫下,SbMT-2基因改造煙草植株的qP值和ФPSII值均高於WT植株,這表明SbMT-2的過表達可以提高植物光合電子傳遞的效率。 在本研究中,轉錄組分析表明,在pHBA脅迫下,PeMPK7基因過表達植株表達的MT-2水平大大高於WT植株。 PeMPK7可能在MT-2基因的上遊發揮作用,並透過調節MT-2的表達提高楊樹的光合效率。這一點應在今後的研究中加以驗證。入射光、環境溫度、微量元素和水分供應的變化對光合作用有很大影響。當入射光水平超過光合作用的吸收能力時,會對植物造成損害,導致光損傷。為了避免光損傷,植物會利用NPQ機制將多余的能量以熱量的形式耗散。NPQ有多種型別,如qE、qH、qZ和qL。qE機制依賴於類囊體蛋白PsbS和黃綠素的可逆光依賴性轉化。受到光照後,質子梯度會在類囊體膜上形成。這引發了PsbS的質子化,並透過啟用中黃素去氧化酶等酶產生玉米黃質,然後將其淬滅。 本研究結果表明,當pHBA濃度低於6mM時,過表達PeMPK7和WT楊樹葉片的NPQ隨pHBA濃度的增加而增加。然而,當pHBA濃度為6mM時,WT植物的NPQ迅速下降,而PeMPK7基因表現植物的NPQ仍呈上升趨勢 (圖9)。這一結果與Li等人的研究結果不一致。 研究者推測,當pHBA濃度高於6mM時,超過了WT楊樹的調節範圍,對PSII反應中心的光合裝置造成嚴重破壞。這導致了PSII反應中心的破壞。結合轉錄組分析結果,PeMPK7過表達植株的PsbS表達量明顯高於WT植株( p <0.05) (圖2B和圖12)。 這表明過表達PeMPK7能增加NPQ和散熱,保護PSII反應中心的光合裝置,並透過調節PsbS的表丹瑞持其正常的光合作用 。這與之前利用VIGS技術在牡丹中瞬時沈默PlMAPK1的研究結果一致;該研究報道基因改造植株的NPQ低於WT植株。因此,研究者推測C組中的MAPK1和MAPK7基因可能在改善NPQ和增加熱耗散以保護PSII反應中心的光合裝置和維持光合作用方面具有功能保護作用。
在本研究中,轉錄組分析顯示PSI和PSII系統中的LHCA1-4、LHCB1-5、petC、psaC、psaK、psaO、psaH、psaL、psbP、psbO和psbR以及Cytb6f中的PetC等基因上調 (圖2A、2B和12)。同時, WGCNA和共表達網路的構建確定了LHCA1、LHCA2、PetA和FaQR基因是光合作用過程中的樞紐基因 (圖5C-E)。 這些結果表明,過表達PeMPK7可透過增強光能吸收和光合電子傳遞能力來提高光合效率。
4 PeMPK7參與pHBA脅迫下光合作用相關基因的TF介導調控
TFs透過與目標基因啟動子中的順式調控元件結合來調控基因表現。鑒於多個光合作用相關基因在轉錄本水平上受到pHBA的調控,研究者認為TFs在pHBA介導的這些基因的表達中發揮著重要作用。 本研究透過構建與光合作用相關的子網路,揭示了TFs與光合作用相關基因之間的潛在調控關系 (圖5A)。
本研究構建的光合作用相關子網路主要包括編碼采光葉綠素(LHC)a/b和PSI/II反應中心色素蛋白的基因以及具有調控關系的潛在TFs。 潛在的TFs屬於WRKY、ZIP、ERF、bHLH和bZIP基因家族。據報道,這些TF的同源物參與了光合作用的調控。例如,植物色素交互作用因子(PIFs)是葉綠素和Car生物合成的負調控因子,植物色素部份抑制PIFs以實作光自營生長。bZIP成員HY5是一種有效的PIF拮抗劑,它能促進光合色素的積累以應對光照。此外,HY5還能控制參與光合作用的其他基因的轉錄,包括核酮糖-1,5-二磷酸羧化酶/氧合酶(RbcS1A)的小亞基。此外,非典型bHLH TF SlPRE5負向調節基因改造番茄葉片中的葉綠素含量,PebHLp5透過調節光合作用正向調節幹旱脅迫響應。因此,光合作用相關子網路中預測的潛在IFs可能參與調控光合作用相關基因。
迄今為止,尚不清楚植物MAPK基因利用哪些TFs來調控pHBA脅迫下的光合作用。 研究者對PeMPK7過表達植株和WT植株中TFs表達水平差異的分析表明,光合作用相關子網路中的ERF3、WRKY1和WRKY33(圖5A)在PeMPK7過表達植株和WT植株中都隨著pHBA濃度的增加而上調 。此外, 在pHBA處理1和3h時,這三個基因在PeMPK7高表達植株中的表達水平明顯高於WT植株(圖5B)。這表明在pHBA脅迫下,PeMPK7透過正向調節WRKY1、WRKY33和ERF3的表達參與光合作用的調控。 在鹽脅迫下,基因改造菊花中的DgWRKY5可透過增加Pn、Gs和Tr來改善光合作用和生長。ERF蛋白(ERE-結合蛋白,ERE BPs)是從煙草中分離出來的GCC盒結合蛋白。先前的研究利用阿拉伯芥蛋白質微陣列確定了阿拉伯芥中作為MAPKs受質的15個ERF家族成員。其中,阿拉伯芥中病原體反應的正調控因子AtERF104透過與MPK6交互作用而穩定。水稻中的OsEREBP1基因可被BWMK1(病原體誘導的MAPK)磷酸化,從而增加其在體外與順式元素的DNA結合活性。這些研究提示了PeMPK7和ERF3之間潛在的蛋白質交互作用。在pHBA脅迫下,PeMPK7透過正向調節WRKY1、WRKY33和ERF3來調控光合作用,這一點還需要利用遺傳學和分子生物學方法進一步驗證。
5 PeMPK7參與了響應pHBA應激的級聯訊號通路
參與植物抗逆的MAPK級聯對於了解MAPK級聯途徑在響應pHBA脅迫時的調控分子機制至關重要。據報道,有幾種MAPK訊號通路參與了植物對生物和非生物脅迫的響應。例如,水稻中的OsMEKK6-OsMAPK5-SERF1訊號通路參與了對鹽脅迫的響應,阿拉伯芥中的AtMEKK1-AtMKK1/AtMKK2-AtMPK4訊號通路參與了對幹旱脅迫和機械損傷的響應。然而,有關MAPK訊號通路參與楊樹脅迫響應調控的研究卻很少。過表達PtMKK4能顯著增強楊樹的抗旱性,並提高過表達PtMKK4的楊樹中PtP5CS、PtSUS3、PtLTP3和PtDREB8的表達水平。研究者之前的研究證明了PeMPK7和PeMKK3之間的交互作用。 本研究表明,除PeMKK3外,PeMPK7還與PeMKK7/10交互作用(圖10),並且PeMKK7和PeMKK10的表達受pHBA的調控。因此,研究者推測PeMKK3/7/10-PeMPK7通路是參與楊樹對pHBA脅迫響應的重要MAPK訊號通路 。然而,這三條途徑的參與還需要透過體外磷酸化實驗進一步驗證。此外, 研究者的轉錄組結果顯示PeMPK7可能調控下遊TFs,即WRKY1、WRKY33和ERF3,這表明PeMKK3/7/10-PeMPK7通路可能透過直接或間接調控WRKY1/WRKY33/ERF3參與了對pHBA脅迫的響應 (圖12)。 鑒於WRKY1/WRKY33/ERF3可能調控光合作用相關基因(圖5A和12),研究者推測PeMKK3/7/10-PeMPK7模組透過調控WRKY1/WRKY33/ERF3來調控光合作用相關基因。 這一假說有待今後利用遺傳學和分子生物學技術進一步驗證。
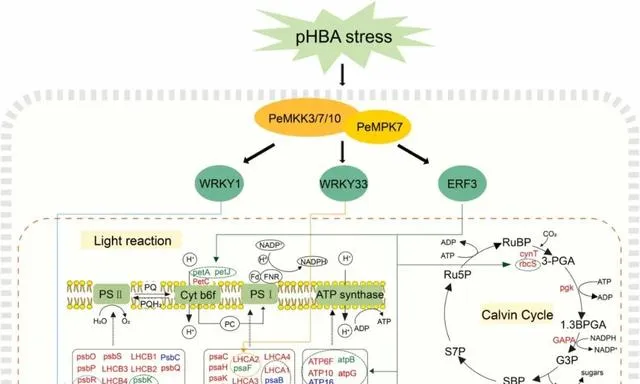
圖12.光合作用相關途徑示意圖。 紅色、藍色和綠色表示與WT植物相比,PeMPK7基因過表達植物在pHBA脅迫後顯著上調、下調和無顯著變化的基因。
結論
總之,
PeMPK7在楊樹中的過表達即使在pHBA脅迫下也能維持高水平的光合作用。本研究透過轉錄組分析、WGCNA和光合作用相關指標的測定,探討了PeMPK7在pHBA脅迫下調控光合作用的分子機制
。
PeMPK7在楊樹中的過表達能在pHBA脅迫早期增加光合作用相關基因的表達。WRKY、ZIP、ERF、bHLH和bZIP基因家族的TFs可能與光合作用相關基因的調控有關。PeMPK7可能在WRKY1、WRKY33和ERF3的上遊發揮作用,並可能透過正向調節它們的表達來促進楊樹的光合作用。
此外,
光合生理指標分析證實,在長期pHBA脅迫下,PeMPK7參與了楊樹光合作用的調控
。此外,
研究者利用酵母雙雜交和拉下實驗證明了PeMPK7和PeMKK7/10之間的交互作用。基於這些結果,研究者推測PeMKK3/7/10-PeMPK7-WRKY1/WRKY33/ERF3通路是參與楊樹對pHBA脅迫響應的重要潛在MAPK訊號通路
。透過構建PeMPK7調控光合作用的通路模型,研究者對MAPK級聯通路調控pHBA脅迫的分子機制有了新的認識。











