編譯:微科盟承樂,編輯:微科盟居居、江舜堯。
微科盟原創微文,歡迎轉發轉載,轉載須註明來源【微生態】公眾號。
導讀
盡管大氣奈米塑膠(NPs)越來越普遍,但對其植物毒性、葉片吸收和在植物中易位的研究仍然有限。本文旨在透過研究番茄葉片暴露於NPs(帶不同電荷)的生理效應以及NPs的葉片吸收和易位來填補這一知識空白。本研究發現,帶正電荷的NPs會引起更顯著的生理效應,包括生長抑制、抗氧化酶活性增加、基因表現和代謝物組成改變,甚至顯著改變了葉際微生物群落的結構和組成。此外,帶不同電荷的NPs表現出不同的葉片吸收和易位,帶正電荷的NPs更多地滲透到葉片中,並在葉肉細胞內均勻分布。此外,葉片吸收的NPs能夠易位到根部。這些發現為了解大氣NPs與作物之間的交互作用提供了重要見解,並表明NPs在作物中的積累可能對農業生產和食品安全產生負面影響。

圖文摘要
論文ID
原名: Toxicity Mechanisms of Nanoplastics on Crop Growth, Interference of Phyllosphere Microbes, and Evidence for Foliar Penetration and Translocation
譯名: 奈米塑膠對作物生長的毒性機制、葉際微生物的幹擾以及葉滲透和易位的證據
期刊: Environmental Science & Technology
IF: 11.4
發表時間: 2023.11
通訊作者: 劉維濤,周啟星,邢寶山
通訊作者單位: 南開大學環境科學與工程學院;美國麻薩諸塞大學
DOI號: 10.1021/acs.est.3c03649
實驗設計

結果與討論
1 NPs暴露的物理-生化效應
本研究表明,葉片暴露於NPs降低了番茄的生長。此外,在PS-SO3H-H和PS-H處理下觀察到萎黃病癥狀和疾病感染(圖1a,f)。暴露於PS-SO3H的番茄植株鮮重沒有顯著下降。然而,當暴露於PS時,植株鮮重顯著降低( p < 0.05),與對照組相比,分別降低41.7%(PS-L)和51.5%(PS-H)(圖1b)。不同水平的NPs暴露對株高的影響有顯著差異( p < 0.05)。在高劑量下,PS對株高的影響與PS-SO3H也有很大的差異( p < 0.05)(圖1b)。除PS-SO3H-L外,NPs顯著降低了葉片的葉綠素含量( p < 0.05)(圖1c)。有趣的是,低劑量NPs(PSSO3H-L和PS-L)暴露下的凈光合速率(Pn)、蒸散速率(Tr)和氣孔導度(Gs)增加,而高劑量NPs(PSSO3H-H和PS-H)暴露下則降低(圖1d)。低劑量NPs對光合作用的刺激和高劑量NPs的抑制與奈米材料「毒物興奮效應」現象中的劑量−反關系相一致。研究發現,低濃度的奈米顆粒可以透過增加葉綠體膜的滲透性來增強葉綠體希爾(Hill)反應活性並加速O2釋放,從而使Ca2+和Cl−更容易進入放氧復合體的中心,從而促進葉綠體的光合磷酸化活性。
葉片暴露於NPs後的光合抑制被認為是影響植物生長的主要機制。NPs暴露會導致O2-和MDA含量顯著增加。雖然SOD、POD、CAT等抗氧化酶的活性增加,但其清除作用並沒有達到維持穩態所需的基線活性氧(ROS)水平。過量的ROS可能會破壞葉綠體結構,抑制光合作用。在暴露於NPs的葉片細胞中,葉綠體結構增大,嗜鋨顆粒增加,證實了上述觀點(圖1e)。高劑量和低劑量NPs的影響之間存在顯著差異,表明番茄抗氧化應激系統對NPs的葉片暴露水平非常敏感。當將阿拉伯芥的根和玉米葉片暴露於NPs時也觀察到了這種現象。對藻類的研究表明,附著在藻類細胞表面的NPs可以阻擋外部光,從而影響光合作用。這種物理效應也可能適用於高等植物。以往的研究發現,CuO和ZnO奈米顆粒可以吸附在植物表面,從而產生遮蔽作用,抑制植物的光合作用。此外,隨後的研究發現,帶正電荷的NPs(F-PS-Np)比帶負電荷的NPs(F-PS-SO3H)在葉片表面吸附得更多。在表面塗層對植物奈米顆粒吸收影響的研究中也發現了類似的現象。這種差異主要歸因於NPs與葉片表面的靜電交互作用不同。帶負電荷的NPs與帶負電荷的細胞壁之間的靜電排斥可導致積累減少,而帶正電荷的NPs由於靜電吸引而大量積累。

圖1.NPs對番茄生長的影響。NPs對植物形態的影響(a);植株鮮重和株高(b)。NPs對光合作用的影響:葉綠素含量(c);光合作用參數(d);葉片形態(f);CK、PS-SO3H-H和PS-H處理組葉片的超微結構圖(e)。(a)和(f)的比例尺分別是8 cm和6 mm。(b)和(c)中的數據以平均值±SD表示(n=3);不同的小寫字母表示不同處理之間差異顯著( p <0.05)。CK:對照組;PS-SO3H-L/PS-L:番茄植株葉片暴露於0.45 μg d−1帶負電荷/正電荷的NPs下21天;PS-SO3H-H/PS-H:番茄植株葉片暴露於4.5 μg d−1帶負電荷/正電荷的NPs下21天。
2 番茄多組學分析
為了研究上述效應的潛在機制,本研究對PS-SO3H-H和PS-H處理組的番茄葉片進行了轉錄組學和代謝組學分析(從此處開始,統一描述為PS-SO3H和PS)。PS對基因調控的作用強於PS-SO3H(圖2a)。為了進一步研究轉錄組變化,本研究用GO和KEGG對DEGs進行了功能註釋。PS-SO3H和PS均上調了MAPK訊號通路,這是由NPs暴露後的細菌性斑點病感染引起的(圖1f)。MAPK級聯反應在感知病原體訊號分子時被啟用,以增強植物對疾病的抗性。此外,NPs還調節植物-病原體的交互作用通路。這一觀察結果與Wang等人的類似結果一致,他們報道了PS-MPs誘導生菜葉片中與抗病通路相關的基因下調。
除上述幾方面外,NPs還可以透過幹擾光合色素和光合位點的產生來影響植物的光合作用。當暴露於PS-SO3H時,類胡蘿蔔素的生物合成過程明顯下調,這影響了葉片的光吸收和抗氧化能力。GO分析也顯示了類似的結果。PS-SO3H顯著影響蛋白激酶活性和蛋白自磷酸化,這在植物抗逆性中起著關鍵作用,且後者是植物色素的重要來源。對於PS,包括葉綠體類囊體膜和質體類囊體膜在內的GO terms受到顯著影響,這是光合作用的光反應位點。核酮糖-1,5-二磷酸羧化酶是碳固定的關鍵酶,其催化核酮糖-1,5-二磷酸生成3-磷酸甘油酸。在本研究中,PS暴露下調了編碼二磷酸核酮糖羧化酶大亞基的 rbcL ,幹擾了無機碳向有機化合物的轉化,從而抑制了番茄植株的光合效率和生物量。
在代謝組學方面,透過主成分分析(PCA)和偏最小平方判別分析(PLS-DA)發現,NPs暴露顯著改變了葉片的代謝產物。差異代謝物(DM)主要包括碳水化合物、胺基酸和次生代謝物(圖2b)。碳水化合物是植物生長發育所必需的能量來源。松二糖、乳糖、果糖和山梨糖含量的降低可能是由於NPs誘導的氧化應激,這會增加與能量代謝相關的糖降解。胺基酸是各種次生代謝產物的前體,在植物生理過程中起著至關重要的作用。絲胺酸和L-谷胺酸參與葉綠素合成,其含量降低會導致葉綠素含量降低(圖1c)。纈胺酸和脯胺酸可作為抗氧化劑和訊號分子參與植物對非生物脅迫的反應。PS暴露顯著降低了纈胺酸和脯胺酸的水平,表明PS誘導了更多的氧化應激。
本研究對轉錄組學和代謝組學進行了相關網路分析,進一步探索番茄的調控網路。許多DEGs和DMs的相關性很強( p < 0.05,R > 0.9)(圖2c)。這些DEGs主要富集在氨酰-tRNA生物合成、丁酸甲酯代謝、丙胺酸、天門冬胺酸和谷胺酸代謝以及谷胱甘肽代謝通路的代謝物中(圖2d)。谷胱甘肽代謝是植物在面臨脅迫時維持關鍵生物過程的機制。此外,氨酰-tRNA由合成酶形成,是蛋白質合成的組成部份。NPs對氨酰-tRNA生物合成的破壞表明,需要關註作物透過調節轉譯過程來應對NP脅迫的作用。
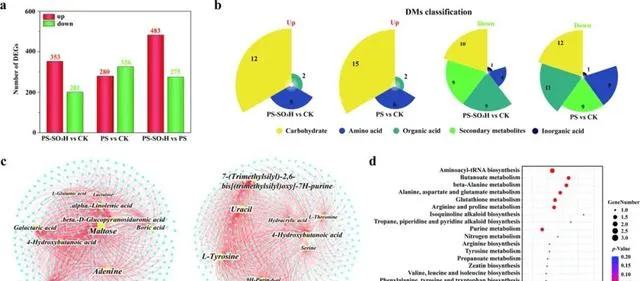
圖2.番茄葉片施用NPs後的轉錄組學和代謝組學分析。番茄葉片的DEGs數量(a)和DMs的分類統計(b)。轉錄組和代謝組變化的關聯分析。不同處理下的前10個DMs和DEGs之間的關系網路(|log2FC|≥2, p ≤0.05)(c);使用氣泡圖顯示共現網路中代謝物的主要代謝通路富集結果(d)。在共現網路中,黃色和青色節點分別代表DMs和DEGs。節點的大小與連線到節點的邊數有關。CK:對照組;PS-SO3H-H/PS-H:番茄植株葉片暴露於4.5 μg d−1帶負電荷/正電荷的NPs下21天。
3 葉際微生物群落的變化
葉際是各種微生物的重要棲息地。葉際為微生物提供營養;微生物有助於植物的生長和發育。當面對外部環境脅迫時,葉際微生物族群可能會發生變化,從而影響它們與宿主的交互作用。
β多樣性分析顯示,各處理間的聚類位置顯著分離,表明NPs對番茄葉際微生物群落結構有顯著影響(圖3a)。α多樣性分析顯示,與對照組相比,NPs暴露提高了Chao、Ace和Shannon指數。特別是,PS暴露更顯著地改變了葉際微生物群落的多樣性,並誘導了特定微生物族群的富集(圖3b)。此外,MNPs也可以改變門水平細菌組成。這種變化通常是由能夠生物降解MNPs的細菌豐度增加而引起的。在本研究中,NP處理組塑膠降解菌(如變形菌門和藍細菌門)的相對豐度增加(圖3c)。在科水平上,芽孢桿菌科在番茄葉際中占優勢,其在對照組中的相對豐度為76.43%,在PS-SO3H和PS暴露後分別降至56.85%和37.06%。芽孢桿菌是公認的有益菌,能夠促進植物生長,抵禦病原菌感染。NPs誘導的芽孢桿菌科相對豐度降低可能是番茄抗病性受到幹擾的原因。
微生物通常會發展出復雜的網路來應對外部幹擾,而核心微生物可以透過微生物之間的交互作用強烈地影響整個微生物群落的結構。叢毛單胞菌科(Comamonadaceae)是一個重要的微生物分類群,已被證明可以有效地控制葉際微生物豐度,其在NPs暴露後顯著增加。本研究分析了番茄葉際微生物的共現網路。NPs處理幹擾了微生物群落的網路復雜性(圖3d)。其中,與對照組相比,NPs暴露後葉際微生物網路的邊數、平均度、聚類系數和網路直徑均顯著增加。有研究表明,葉際微生物透過形成生物膜增加了MNPs在葉片表面的聚集。相應地,NPs在植物表面的沈積為微生物提供了新的生態席位,從而為葉際微生物交互作用提供了機會。這有助於解釋本研究中NPs增強了細菌之間的關系,並加強了微生物群落的共現模式。
植物葉際微生物交互作用在維持植物健康和抑制病原菌過度生長方面發揮著重要作用。它們可能受到多種生物因素(如葉片代謝物和植食性昆蟲)和非生物因素(如氣候因子)的影響。本研究還發現,大量的DMs與番茄葉際細菌門之間存在顯著的相關性。含碳和含氮營養物質的可用性是限制附生微生物定植的主要因素。暴露於NPs後,番茄葉片代謝物中碳水化合物和胺基酸組分顯著增加。以往的研究表明,葉片養分與葉際微生物群落的穩定性呈正相關。這與本研究結果相似,表明NPs可以透過改變植物代謝物來促進新的優勢微生物群落的形成,並形成葉際微生物群落。
利用PICRUSt預測葉際微生物功能的變化。KEGG一級通路中富集三種通路(代謝、細胞過程和環境資訊處理)。在二級通路中,有5條通路發生了顯著改變,其中4條是代謝通路。因此,葉際微生物的代謝也受到了幹擾。此外,PS暴露顯著增加了細胞生長與死亡通路的豐度,從59.93%增加到69.58%。葉際微生物生理過程的提升可能是微生物網路高度復雜的原因。

圖3.葉際微生物群落對NPs的響應。NMDS分析(a),對照組和NPs處理之間細菌α多樣性(Chao、Ace和Shannon指數)比較(b)。細菌門和細菌科的相對豐度(c)。NPs對細菌群落共現模式的影響(d)。節點的大小根據節點的度進行縮放,並根據模組對節點進行著色。(b)中的數據以平均值±SD表示(n=3)。* p <0.05,** p <0.01。CK:對照組;PS-SO3H/PS:番茄植株葉片暴露於4.5 μg d−1帶負電荷/正電荷的NPs下21天。
4 電荷控制NPs內化進入葉片
為了證明番茄葉片對NPs的吸附和吸收,本研究使用CLSM研究了NPs的轉移效率和分布。根據熒光NPs穩定性測試結果,F-PS-SO3H和F-PS-Np NPs在模擬植物溶液和原生質體細胞液中24 h內均未檢測到熒光釋放。沖洗後,在4 h時,葉片表面的F-PS-Np熒光強度強於F-PS-SO3H,表明葉片表面對帶正電荷的NPs的吸附能力更強。由於植物細胞壁帶負電荷,這一發現是符合預期的。此外,本研究發現PS-SO3H與葉片的接觸角高達105.80 ± 0.47,導致液滴呈球形,容易遺失。PS-Np與葉片的接觸角為93.96 ± 0.20,液滴呈扁球形,更容易吸附在葉片上,並可快速擴散。這導致F-PS-Np在葉片表面呈均勻分布。同時,F-PS-SO3H大量聚集,進一步阻礙了NPs的內化。
本研究透過CLSM影像和3D視訊顯示了不同時間NPs在葉肉中的分布和傳遞情況。在CLSM影像中,觀察到了更多的F-PS-Np,表明帶正電荷的NPs比帶負電荷的NPs更能有效地進入葉肉。隨著時間的推移,兩種NPs的轉移效率都有所提高(圖4)。F-PS-Np和F-PS-SO3H的分布也有顯著差異。F-PS-Np均勻分布並被轉移到葉肉細胞中。相比之下,F-PS-SO3H只有在暴露時間超過24 h後才會少量進入細胞內部(圖4a)。
氣孔和表皮被認為是葉片吸收NPs的兩個主要途徑。本研究發現,葉片可以透過這兩種途徑吸收帶不同電荷的NPs。NPs可以透過質外體(胞外)和共質體(胞內)途徑在葉片內易位。本研究發現,帶正電荷的NPs在葉綠體中的轉運途徑是共質體,而帶負電荷的NPs則透過質外體途徑轉運(圖5)。後者更容易理解,因為雙子葉植物的葉子同時包含薄膜狀和海綿狀的葉細胞。這些細胞排列稀疏,形成了較大的胞外空間,但NPs從細胞外空間進入葉肉細胞時會遇到細胞壁和細胞膜等屏障。
植物細胞壁被認為是阻擋奈米材料的主要屏障,其標準平均孔徑小於10 nm,遠遠小於本研究中的NPs。雖然奈米顆粒的吸收和易位受到尺寸排斥的限制,但與本研究結果相似,據報道高達36−50 nm的工程奈米顆粒(ENPs)可以被內化和易位。尺寸排斥是動態的,並直接受到鈣、蛋白質、病毒或ENPs的影響。常見的ENPs(如SiO2NPs和TiO2 NPs)可以誘導細胞壁形成新的或更大的孔隙,從而改變細胞壁結構,促進較大ENPs的內化。與這些ENPs相比,本研究中使用的NPs更柔軟。本研究觀察到被植物內化的NPs發生了變形(圖7c),這有利於NPs的吸收和易位。Li等人也報道了類似的發現。
由於發現NPs已經進入植物細胞,本文研究了原生質體(無細胞壁)對NPs的攝取。本研究在健康的原生質體中發現了與完整葉片相同的積累模式,其中F-PS-SO3H聚集在原生質體周圍,F-PS-Np在原生質體中聚集。與本研究結果相似,有報道稱,帶正電荷的顆粒比帶負電荷的顆粒更容易被細胞吸附並透過內吞作用穿過細胞膜。然而,也有報道稱,軟塑膠顆粒的表面正電荷抑制了它們穿透植物細胞膜的能力。由於軟塑膠的特異性,植物細胞可能透過不同於硬塑膠的途徑來吸收它們。例如ZnO和Au等硬顆粒可以進入原生質體,而與電荷無關。與帶負電荷的粒子相比,帶正電荷的粒子被快速內化,這與本研究結果相似。易位的差異可能與粒子的zeta電位有關。據報道,隨著奈米顆粒zeta電位的增加,脂質雙分子層誘導的電位下降也隨之增加,這將導致膜張力的增加,從而導致奈米顆粒更強的易位。在本研究中,F-PS-Np的zeta電位絕對值高於F-PS-SO3H,表明這一因素可能會影響NPs的易位效率。
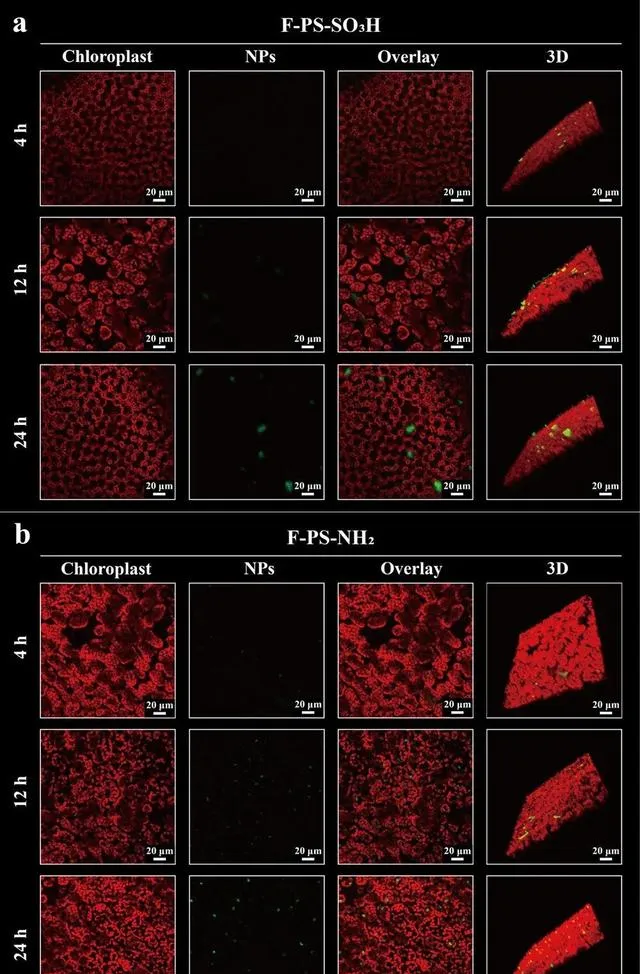
圖4.葉片中NPs的易位和分布。4 h、12 h和24 h時葉肉中F-PS-SO3H (a)和F-PS-Np (b) NPs熒光強度的CLSM影像。番茄植株葉片暴露於F-PS-SO3H或F-PS-Np NPs(約2μg NPs/葉)。綠色是NP粒子,紅色是葉綠體。
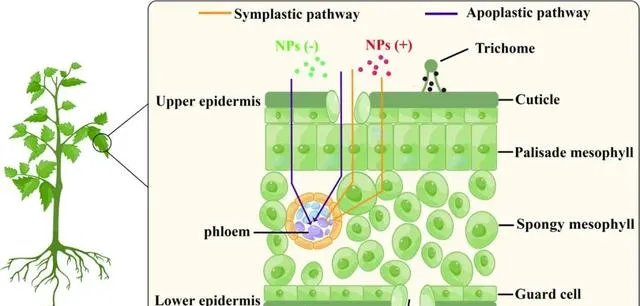
圖5.番茄葉片中帶不同電荷的NPs的易位途徑及其分布。NPs透過氣孔和/或表皮途徑被吸收到葉片中,然後透過質外體途徑(紫色線)在細胞外空間和細胞壁之間轉移,或透過共質體途徑(橙色線)進入葉肉細胞。表面電荷的差異影響NPs的易位途徑。NP(−):帶負電荷的NPs;NP(+):帶正電荷的NPs。
5 NPs的葉片-根部易位及分布
為了進一步證實NPs的吸收和易位,本研究使用直徑為70 nm的聚苯乙烯奈米塑膠摻雜銪螯合物Eu-β-diketonate (PS-Eu),以定量番茄不同組織中的NPs。對於維管組織,韌皮部負責將光合產物從葉片長距離運輸到其他組織。本研究發現,被葉片吸收的PS-Eu可以發生易位,但其易位能力有限。在第7天,內化到葉片中的PS-Eu只有3.51 ± 0.092%轉移到根部,且隨著施用劑量和時間的增加沒有顯著變化(圖6b)。透過掃描電子顯微鏡(SEM)也觀察到了同樣的現象(圖7)。很多PS-Eu顆粒在番茄葉片中聚整合簇(圖6c,7a-f)。在根部只觀察到少量的PS-Eu,其呈現為單獨分散的顆粒或是聚整合簇(圖6c,7g-l)。
造成這種現象的原因有很多,其中NPs的大小和聚集狀態被認為是影響吸收和易位的主要因素。葉片吸收的大小限制可能取決於植物型別和NP特性等因素,因此確切的限值仍不清楚。一項研究表明,100 nm的PSNPs也可以被葉片吸收並轉運到植物根部,其粒徑比本研究選擇的PS-Eu NPs大得多。NPs的聚集是抑制還是減少葉片吸收尚不清楚,但有報道稱聚集的NPs被保留在葉片表面,這在本研究中也有發現。被葉片吸收的NPs在到丹瑞管組織之前必須穿過幾個屏障,包括表皮、質外體或共質體,這些都將進一步阻止NPs在葉片內的運輸。施用於葉片的NPs到達韌皮部後,可透過由加寬的原生質絲組成的篩管轉運到植物的其他部位。NPs在韌皮部的易位過程中會遇到光合產物、螯合分子和不同的pH梯度。在這一過程中,NPs可能聚集、溶解或形成蛋白冠。因此,除了大小限制外,NPs的韌皮部轉運非常復雜,其具體機制尚不清楚,仍有待進一步探索。
本研究在405 nm激發下,測定了PS-Eu處理7 d的番茄幼苗。在番茄根、莖和葉中觀察到藍色熒光(Eu的自身熒光)。本研究還發現,藍色熒光主要集中在莖的韌皮部,這是NPs在韌皮部運輸的直接證據。毛狀體被認為是工程奈米顆粒進入葉片的可能途徑。有趣的是,本研究也在葉片的毛狀體中發現了PS-Eu。然而,這些NPs是被毛狀體直接內化,還是被表皮內化後轉移到毛狀體內,仍有待進一步探索。此外,本研究在番茄莖的毛狀體內部和外部都發現了PS-Eu,即使它們沒有直接暴露於PS-Eu。PS-Eu很可能透過毛狀體特定部位的組織間隙被擠壓出來,這可能是植物透過自身排泄來解毒的主要機制。
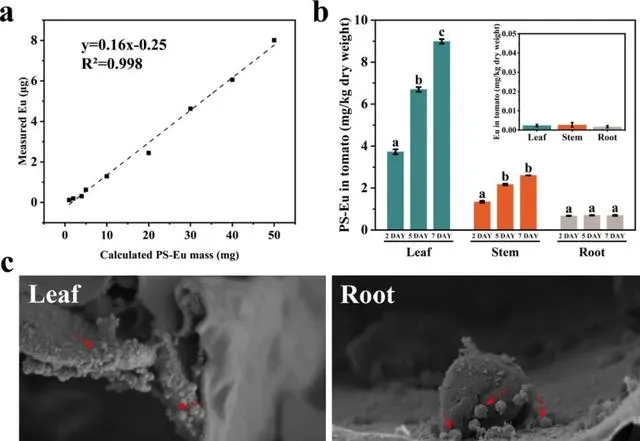
圖6. PS-Eu在番茄中的分布。PS-Eu計算品質與Eu之間的相關性(a)。2、5和7天時PS-Eu NPs在番茄葉片、莖和根中的生物累積(b)。右上方的圖顯示了未暴露PS-Eu植株不同組織中Eu的含量。(a)和(b)中的數據以平均值±SD表示(n=3)。不同字母表示處理天數之間存在顯著差異( p <0.05)。PS-Eu在番茄葉片和根中的分布(c)。番茄植株葉片連續7天暴露於2 μg PS-Eu NPs/leaf。實心箭頭表示PS-Eu。

圖7. PS-Eu在番茄葉片(a-f)和根部(g-l)分布的SEM影像。番茄植株葉片連續7天暴露於2 μg PS-Eu NPs/leaf。每個虛線箭頭表示紅框內區域的放大檢視。實心箭頭表示PS-Eu粒子(c,f,i和l)。
結論
綜上所述,NPs可以進入番茄葉片,但帶不同電荷的NPs在葉片吸收和分布方面存在顯著差異。正電荷有利於NPs進入葉片並在葉片內均勻分布。NPs的細胞攝取也觀察到類似的結果。NPs在番茄組織中的積累和分布對植物生長有不同的影響,帶正電荷的NPs比帶負電荷的NPs對植株生長的抑制更嚴重。此外,積累在葉片中的NPs可以透過韌皮部運輸到植物根部。未來需要註意的是,在葉片中積累的NPs是否會轉移到可食用的果實部份,從而對人類健康造成潛在危害。NPs在韌皮部的轉運機制尚不清楚。有必要利用蛋白質組學等方法來探索可能參與NPs韌皮部運輸的重要蛋白質。總體而言,本研究有助於更好地理解NPs與作物葉片的交互作用,這有助於了解NPs的歸趨和影響。
然而,受檢測技術和取樣困難的限制,只有少量研究調查了大氣中NPs的含量、組成和形態。為了獲得更真實的環境NPs水平的影響,隨後應嘗試收集灰塵和蔬菜樣本,並確定NPs的數量和型別,以進行進一步的研究。此外,NPs還會經歷風化、分解等過程,從而導致這些NPs的理化性質發生變化。因此,有必要在更實際的環境下探討風化NPs對作物生長和營養品質的影響。











