
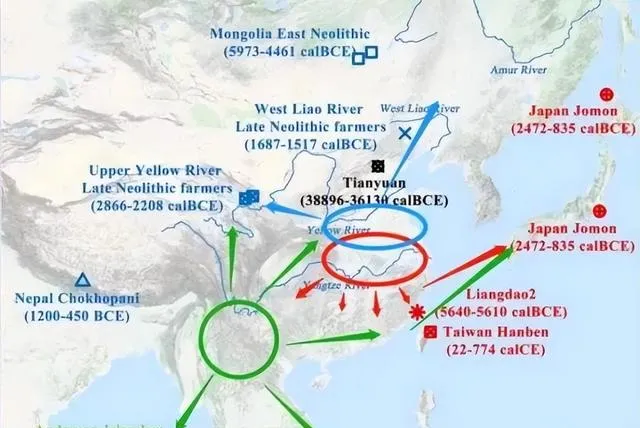
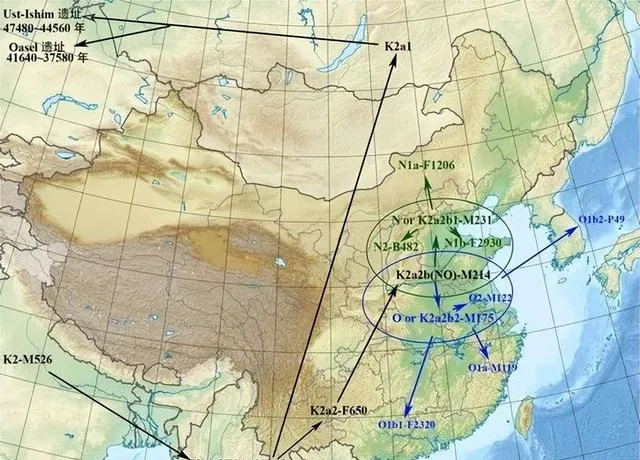
关于东亚父系NO起源问题,多数人认同西南起源说,但相当部分人认为西北/北方起源说。双方各持一辞,似各有道理,但事实真的如此吗?随着随着古DNA提取及测序技术不断完善,南北各地古DNA数据量越来越大,进一步深化了我们对祖先起源的认识。现有的古DNA数据,与西南起源说、西域起源说有很大的出入。根据考古调查报告,新石器时代早期中期华北地区及其山东一带发现了很多N-M231,晚期开始出现了O-M175,南方新石器中期至末期出现了很多O-M175、但N-M231极为稀缺。已知K2a2b(NO)-M214有两大分支:N-M231与O-M175。如果哪些地区N-M231与O-M175交替共存,下游分支齐全,那么最可能是NO-M214发源地,向周边地区扩张。
在黄河中下游,N-M231的下游分支丰富多样,包括N1-CTS3750与N2-B482。其中N1-CTS3750衍生了两大分支N1a-F1206与N1b-F2930。N2-B482集中分布于中国华北、东欧。
N1a-F1206:东欧、西伯利亚、中国北方,但N1b-F2930稀缺。
N1b-F2930:中国东部、藏缅语族、中南半岛,但N1a-F1206稀缺。
N-M231的下游分支(N1a、N1b、N2)区域交集,结果为黄河中下游,可以推测黄河中下游最可能是N-M231发源地,粟作农业文化人群与N-M231具有密切相关性。
O-M175有两大分支:O1-F265与O2-M122。其中,O1-F265衍生了两大分支:O1a-M119与O1b-M268。
O1a-M119:中国南方、东南亚。
O1b-M268衍生了两大分支:东南亚O1b1-F2320与日韩O1b2-P49
O2-M122衍生了两大分支:东亚/太平洋O2a-M324与华东O2b-F742
O-M175的下游分支(O1a-M119、O1b-M268、O2-M122)区域交集,结果为东亚区域,可以推测O-M175最早分化地点最可能是中国东部,稻作农业文化人群与O-M175具有密切相关性。
不难看出:在黄淮海平原(华北与淮河之间),N-M231与O-M175交替共存,下游分支齐全,那么最可能是NO-M214发源地。

距今六千~五千年的鲁北大汶口文化傅家遗址男性则有多样化的Y染色体序列,实际测到K*(×O)有一例、N为两例、O为一例。同期河北红山文化姜家梁遗址全是12例N。由于鲁北与河北为邻,,必有人员流动,出现了N、O并存的现象,清晰表明大汶口文化北上扩张的过程,进一步证明新石器时代山东地区N-M231与O-M175交替共存,最可能是NO-M214发源地。
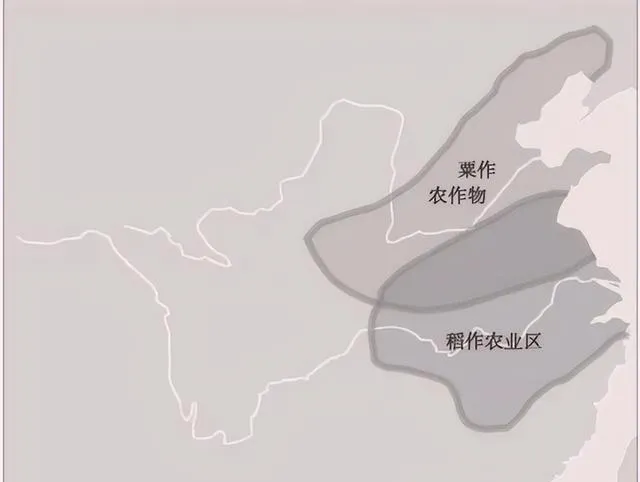
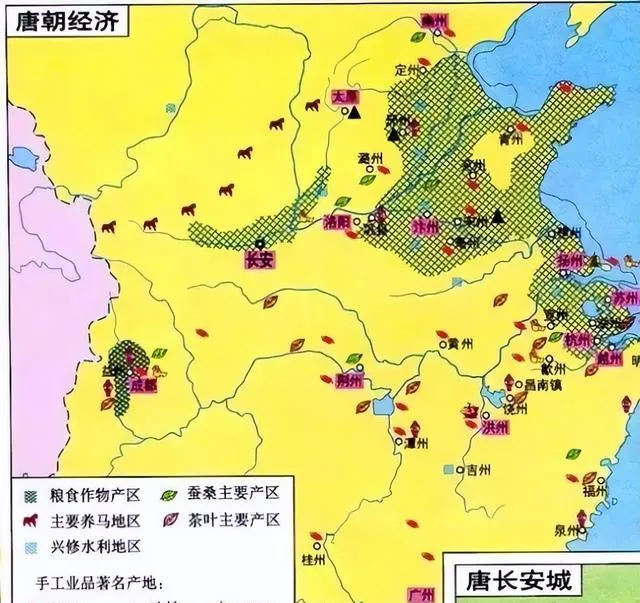
此图出自农业部。古代主要粮食作物产区地几乎都在黄河中下流域,而西北地区、北方地区都是游牧民族的主要养马地区,对于东方农业文明构成严重的威胁。
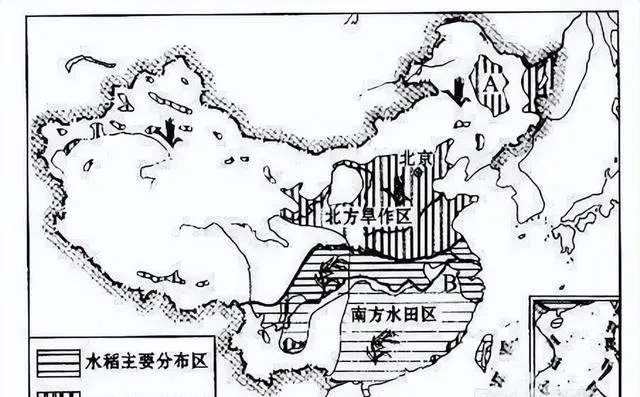
N系人群率先到达黄河流域北岸,开创了粟作农业,但生产规模不大,至今还没有成为中国人的主粮,N系人群渐渐地退出历史舞台,但留下一定的文化遗产。
O2-M122早在3万年前就已到达东亚,掌握了农业种植技术,在来东亚之后才开始其新石器时代的扩张,与中国农业的东部本土起源是一致的。总之,O2-M122强烈人口扩张的Y染色体支系在中国东部的广泛分布,由于中国东部大平原属半湿润、半干旱气候地区,当初从事稻粟混作,后来引进了小麦,致力于在中原地区推广小麦种植,奠定了黄河农业文明的基础,小麦最终称为北方汉人的主粮。东南亚O1a-M119和O1b1a1-M95长期生活在热带森林从事半农半猎,远不如黄河农业文明高度发达。湖南、浙江一带最早出现水稻,但生产规模不大,也不是商周秦汉的主粮。自从汉代统一南方,才开始开拓南方农业,稻栽培已具相当规模。自从宋代以来,水稻慢慢变成了多数中国人的主粮。

Ust-Ishim人(乌斯特-伊斯姆人是K2A)是在俄罗斯西伯利亚西北地区发现的4.5万年前的Y染色体为k2a的人类遗骨。K2a 或者说pre NO是NO的祖形(未分化),和其他的K2支系相比分化较早, 其他的K2支系包括K2b、K2c、K2d、K2e。K2a 最早与其他K2集团分离,在印度巴基斯坦地带向北迁徙, 其他K2则继续挺进南亚、东南亚、澳大利亚。所以,东南亚K2多样性很高,推测K2a-CTS11667最可能起源于南线,估五万年前到达东亚沿海,没有继承北欧亚ANE人群的一丁点常染,但是继承了田园洞人的东欧亚常染。田园洞人或者Ust-Ishim人是东欧亚、南欧亚、巴布亚澳洲共同的一个祖先,并且现代东亚NO人群不含ANE常染成分,否定了NO西北/北方起源说。


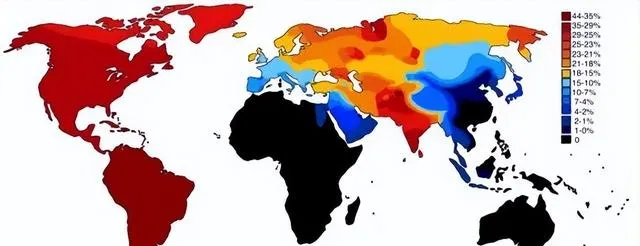
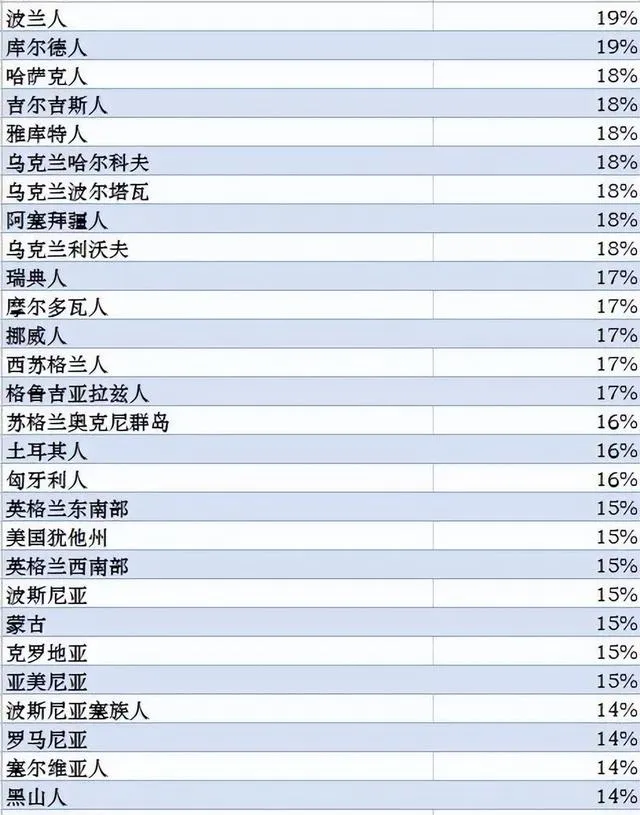
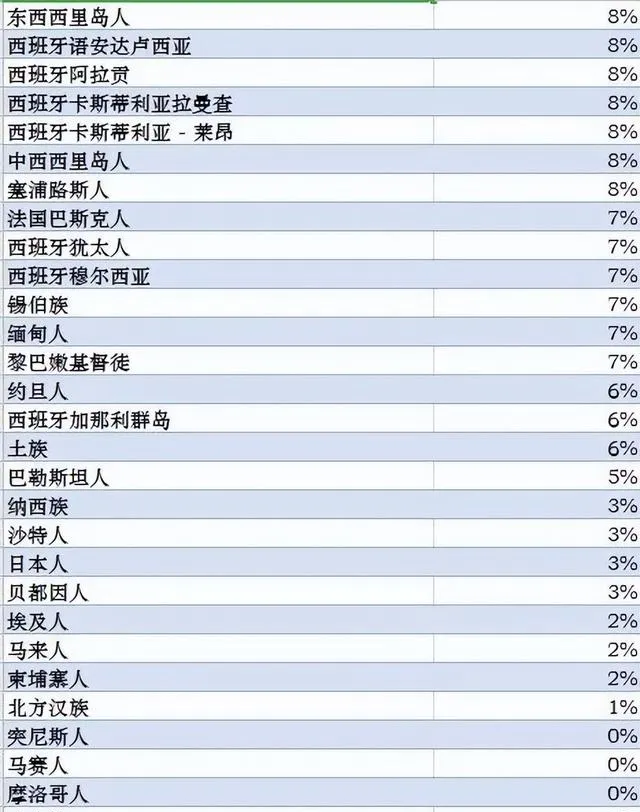
世界各族ANE血统比例分布图,ANE古北欧亚人是大概两万年前中石器时代分布在欧亚大陆北部的一个人群,可以解析为70%的西欧亚和30%的东欧亚血统的混合。参考此数据制作而成。




首先要了解一下现代欧洲人形成的过程。7万年前智人走出非洲,在中东混入了少量尼安德特人基因,然后分离成东欧亚人(East Eurasian)、西欧亚人(West Eurasian)这两大分支。有一支没混入尼安德特人基因的人群叫Basal Eurasian留在了中东和北非。由于没有混入尼人基因,Basal Eurasian比较接近撒哈拉以南的非洲人。
东欧亚人东迁后分离成形成东亚人种、大洋洲土著人种。
而西欧亚人迁徙到欧洲,形成西欧亚狩猎采集者(WHG)。另一种西欧亚人迁徙到西伯利亚,混入了30%~20%的东欧亚基因形成古北欧亚人(ANE),之所以现代西伯利亚土著不是高加索人种是因为后来东亚人种不断北上迁徙稀释当地ANE血统的结果。ANE对后期草原牧民古印欧人有很大的血统贡献,后来印欧人大规模迁徙到欧洲。新石器时代一支西欧亚人和中东的Basal Eurasian人混合形成中东新石器时代农民(EEF),EEF也大规模迁徙到欧洲,并带来农业种植技术。
现代欧洲人正是西欧亚狩猎采集者(WHG)、古北欧亚人(ANE)、Basal Eurasian这三种遗传差异较大的人群混合的结果。根据对欧洲古人类基因的检测,现西欧亚狩猎采集者WHG人群大多同时具有深色皮肤、蓝眼睛、黑头发的特征,但遗传特征接近现代北欧人。

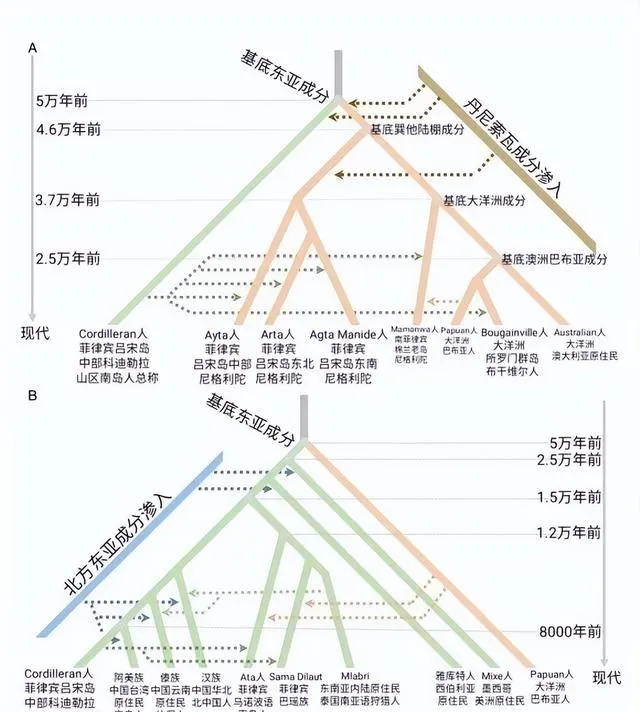

付巧妹团队研究观察到,现今的东亚人群,不管是来自北方还是南方,都同时混合有古北方人群成分(以黄河沿岸的山东人群为代表)和古南方人群成分(以福建与台湾海峡岛屿人群为代表),但古北方人群成分影响更为广泛(图2C和图3C)。而与福建奇和洞人相关的古南方人群成分在中国南方大陆大量下降(图3C和3F)。该结果表明人群的迁移,包括新石器时代南北方人群之间的互动融合,以及 新石器时代以后黄河以北人群向南的大量迁移,基本构成了现今东亚南北方人群的基因组成。
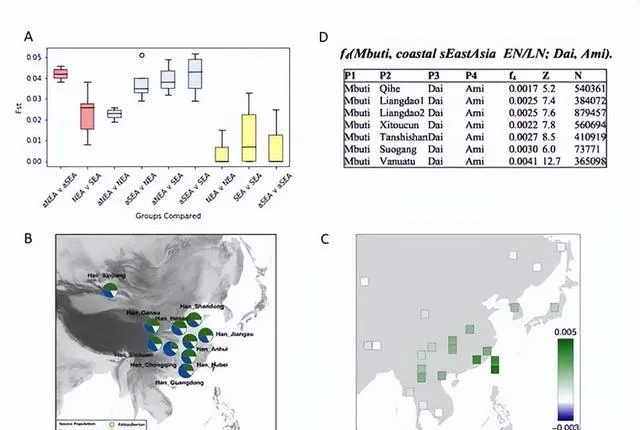
(A)不同人群的遗传分化指数分布图,指数越高表示相关人群的分化程度越高,遗传差异越大,该图表明新石器时代南北方人群的的遗传差异远高于现今东亚南北方人群。(B)现代汉族人群祖源成分与地理分布图。 现今中国南北方的汉族人群,不管是在南方还是北方,均显示混合有东亚古南方人群成分、古北方人群成分、古西伯利亚人成分。其中,现代汉族人群里携有古北方人群成分比例最高的群体显示位于山东,这里也正是本研究古北方人群的代表个体样本的遗址所在地,由此反映出山东地区人群自新石器时代以来的遗传连续性。 (C)现今东亚人群与古南方人群、北方达斡尔族人群的遗传关系图,绿色表示与古南方人群的关系更近,蓝色表示与达斡尔族人群更近。(D)古南方人群与现今傣族人群、台湾岛阿美族人群的遗传关系的f4分析图,Z>3 表示与阿美族人群关系更近。

复旦伦敦新论文中的截图译文:(图4B)进行的成对遗传同质性的检验结果证实,汉代个体(受降城汉军烈士、大槽子汉墓)与新石器早期山东个体的遗传同质性高于黄河内陆(仰韶组和齐家组),汉代个体在PCA(图1A)和qpAdm(图3B)中的位置更接近于新石器时代的黄河先民。这表明,黄河内陆与沿海的混合在北方汉人的形成中起着重要作用。
这个树图出自2021年3月发表的【全基因组见证河西走廊人群混合史】论文,明确指出:九千年前山东小高遗址是100%东亚人后裔。北方粟作农人是由68%东亚成分和 32%类Onge成分混合的,仰韶文化汪沟遗址是北方粟作农人的直属后裔。南方稻作农人是由77%类Onge成分何23%东亚成分混合的,台湾汉本遗址是南方稻作农人的直属后裔。甘肃兰州汉族是由97%北方粟作农人和3%南方稻作农人混合的。
研究发现,广西地区距今约1500年至500年的古人群与距今6000年的史前古老人群(包括广西隆林人、独山人和宝剑山人)完全不同,这意味着广西特有的以隆林人为代表的相关古老东亚人群没有任何基因贡献,远古广西人彻底灭绝了。古代广西人群更替,见证古东亚北方人群的扩张之道。九千年前古山东常染(古东亚北方人群),最终成为北汉血统主体。这一事实清楚告诉:K2a祖先五万年前到达东亚沿海后所形成的东欧亚常染成分,作为东亚人种的奠基者,与后期南方的南欧亚成分(澳棕类型、和平文化人群)完全不同。
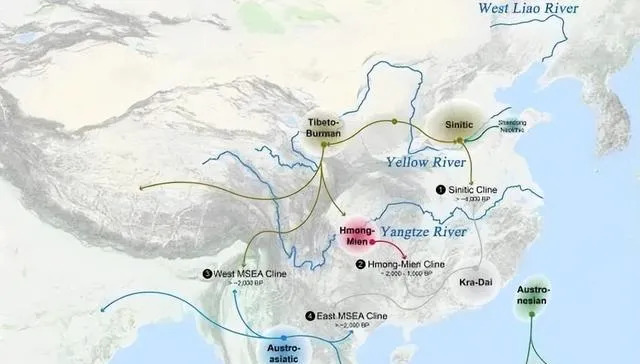
下面的祖先迁徙路线图,又简单又易懂。
原始东亚人(东亚农业人群)起源于黄淮海大平原,向西北/西南、华北/北亚、华南/东南亚、东北亚/日韩迁徙扩散,与当地土著游牧人群或渔猎人群混血,形成了亚洲各族。
首先要了解一下现代欧洲人形成的过程。7万年前智人走出非洲,在中东混入了少量尼安德特人基因,然后分离成东欧亚人(East Eurasian)、西欧亚人(West Eurasian)这两大分支。有一支没混入尼安德特人基因的人群叫Basal Eurasian留在了中东和北非。由于没有混入尼人基因,Basal Eurasian比较接近撒哈拉以南的非洲人。
东欧亚人东迁后分离成形成东亚人种、大洋洲土著人种。
而西欧亚人迁徙到欧洲,形成西欧亚狩猎采集者(WHG)。另一种西欧亚人迁徙到西伯利亚,混入了30%~20%的东欧亚基因形成古北欧亚人(ANE),之所以现代西伯利亚土著不是高加索人种是因为后来东亚人种不断北上迁徙稀释当地ANE血统的结果。ANE对后期草原牧民古印欧人有很大的血统贡献,后来印欧人大规模迁徙到欧洲。新石器时代一支西欧亚人和中东的Basal Eurasian人混合形成中东新石器时代农民(EEF),EEF也大规模迁徙到欧洲,并带来农业种植技术。
现代欧洲人正是西欧亚狩猎采集者(WHG)、古北欧亚人(ANE)、Basal Eurasian这三种遗传差异较大的人群混合的结果。根据对欧洲古人类基因的检测,现西欧亚狩猎采集者WHG人群大多同时具有深色皮肤、蓝眼睛、黑头发的特征,但遗传特征接近现代北欧人。
付巧妹团队研究观察到,现今的东亚人群,不管是来自北方还是南方,都同时混合有古北方人群成分(以黄河沿岸的山东人群为代表)和古南方人群成分(以福建与台湾海峡岛屿人群为代表),但古北方人群成分影响更为广泛(图2C和图3C)。而与福建奇和洞人相关的古南方人群成分在中国南方大陆大量下降(图3C和3F)。该结果表明人群的迁移,包括新石器时代南北方人群之间的互动融合,以及新石器时代以后黄河以北人群向南的大量迁移,基本构成了现今东亚南北方人群的基因组成。
(A)不同人群的遗传分化指数分布图,指数越高表示相关人群的分化程度越高,遗传差异越大,该图表明新石器时代南北方人群的的遗传差异远高于现今东亚南北方人群。(B)现代汉族人群祖源成分与地理分布图。现今中国南北方的汉族人群,不管是在南方还是北方,均显示混合有东亚古南方人群成分、古北方人群成分、古西伯利亚人成分。其中,现代汉族人群里携有古北方人群成分比例最高的群体显示位于山东,这里也正是本研究古北方人群的代表个体样本的遗址所在地,由此反映出山东地区人群自新石器时代以来的遗传连续性。(C)现今东亚人群与古南方人群、北方达斡尔族人群的遗传关系图,绿色表示与古南方人群的关系更近,蓝色表示与达斡尔族人群更近。(D)古南方人群与现今傣族人群、台湾岛阿美族人群的遗传关系的f4分析图,Z>3 表示与阿美族人群关系更近。
复旦伦敦新论文中的截图译文:(图4B)进行的成对遗传同质性的检验结果证实,汉代个体(受降城汉军烈士、大槽子汉墓)与新石器早期山东个体的遗传同质性高于黄河内陆(仰韶组和齐家组),汉代个体在PCA(图1A)和qpAdm(图3B)中的位置更接近于新石器时代的黄河先民。这表明,黄河内陆与沿海的混合在北方汉人的形成中起着重要作用。
这个树图出自2021年3月发表的【全基因组见证河西走廊人群混合史】论文,明确指出:九千年前山东小高遗址是100%东亚人后裔。北方粟作农人是由68%东亚成分和 32%类Onge成分混合的,仰韶文化汪沟遗址是北方粟作农人的直属后裔。南方稻作农人是由77%类Onge成分何23%东亚成分混合的,台湾汉本遗址是南方稻作农人的直属后裔。甘肃兰州汉族是由97%北方粟作农人和3%南方稻作农人混合的。
付巧妹团队研究发现,广西地区距今约1500年至500年的古人群与距今6000年的史前古老人群(包括广西隆林人、独山人和宝剑山人)完全不同,这意味着广西特有的以隆林人为代表的相关古老东亚人群没有任何基因贡献,远古广西人彻底灭绝了。古代广西人群更替,见证古东亚北方人群的扩张之道。九千年前古山东常染(古东亚北方人群),最终成为北汉血统主体。这一事实清楚告诉:K2a祖先五万年前到达东亚沿海后所形成的东欧亚常染成分,作为东亚人种的奠基者,与后期南方的南欧亚成分(澳棕类型、和平文化人群)完全不同。











