编译:微科盟未知,编辑:微科盟Emma、江舜尧。
微科盟原创微文,欢迎转发转载。
导读
转录组、蛋白质组和代谢组构成了植物基因型稳定性的柔性物质基础,使其能够产生随环境变化而动态变化的表型。将转录组、蛋白质组和代谢组嵌入植物代谢途径通常需要进行复杂且异位的生物技术实验。由质谱成像(mass spectrometry imaging,MSI)主导的空间多组学技术通过相关转录因子、功能性蛋白质和代谢物在通路中的空间关联,提供了最直观的原位证据链。这种空间证据可以与空间、时间、植物形态、组织器官相关联,并为植物表型变化与其大分子或小分子组分之间的动态关系提供最直接的生理证据。本文介绍了MSI如何实现高通量原位空间代谢数据采集的仪器基础以及影响成像质量的因素。接下来的内容涵盖了过去3-4年的空间证据,总结了特定的植物分子生理应用,包括植物天然产物的空间生物合成途径、植物根与外部环境之间的物质交换、植物在非生物胁迫下的生理抵抗、植物在生物胁迫下的化学防御以及植物与微生物之间的相互作用。最后,讨论了将空间多组学与表型技术结合,以实现未来植物生理研究中基因型-转录组-蛋白质组-代谢组-表型的时空一致性的可能性,并探讨了从空间数据中挖掘植物生理信息的困难。
论文ID
原名: Mass spectrometry imaging: an emerging technology in plant molecular physiology
译名: 质谱成像:植物分子生理学的新兴技术
期刊: Phytochemistry Reviews
IF: 7.7
发表时间: 2023.11
通讯作者: 李林楠,徐红
通讯作者单位: 上海中医药大学中药研究所
结果
1 介绍
为了探究植物中稳定基因型与动态表型之间的适应性代谢系统,将转录组、蛋白质组和代谢组相结合的多组学模型,已成为一种流行的技术。多组学提供的多个大型数据集可以从基元水平、方法学水平和数学水平上整合和提取有效信息。此外,多组学技术在探索植物在环境中动态表型与稳定基因型之间的调控方面取得了重大突破。然而,每种植物都具有动态的整体内部代谢系统,使其能够灵活改变表型以维持稳态。地球上超过391000种植物产生数百万种代谢产物。从转录组、蛋白质组和代谢组的大型数据库的有效整合仍然是多组学发展的关键问题。近年来,一些微小分子的空间定位方法已经推动了与时间、空间和植物形态等维度相关的新属性的发展,以研究植物基因型和表型之间的物质代谢。
在植物生理学的发展中,视觉信息通常被视为最直观的补充证据。此外,自1920年代以来,对植物结构可视化的探索方法一直深入到微观水平(图1a)。植物可视化已经发展到可以在组织、器官、细胞,甚至亚细胞水平精确定位植物代谢途径中的转录组、蛋白质组和代谢组的程度。质谱成像(MSI)起源于物理学和材料科学,通过开发能够直接从样本中提取分析物的离子源,如脱附电喷雾电离(desorption electricity spray ionization,DESI)和基质辅助激光解吸/电离(matrix-assisted laser desorption/ionization,MALDI)。然后,MSI应用于医学和生物学领域,在显微镜和免疫组化技术的帮助下,逐渐成为生物组织中蛋白质和肽的空间分析的热门方法。近年来,MSI已广泛用于植物和天然产物的空间代谢组学。根据最近的文献,生物组织的MSI主要用于药代动力学,以研究药物成分在实验动物中的吸收、组织传递和代谢转化的过程。这项研究的关键挑战是排除来自动物内源成分的显著干扰。相比之下,植物MSI倾向于对植物代谢的组织进行全面的空间表征;在这里,困难在于组织切片以及植物代谢的多样性和复杂性。基于MSI,简单处理的植物可用于收集化学成分的原位空间分布数据。通过质量对电荷比(m/z)鉴定的每个代谢物离子可以通过颜色差异的点阵矩阵来表征在组织中的丰度分布,从而可以同时实现多个小分子的组织定位(图1b)。
与其他空间表征方法(如显微检查和免疫组织化学)相比, MSI具有许多优势,包括无可比拟的便利性、高通量和广泛适用性,可用于研究植物代谢多样性。除了生物样本中的蛋白质、脂质、核酸、糖类和激素外,碱性化合物、类黄酮、寡糖和植物激素的定性和定量分析也在基于MSI的空间代谢组学的应用范围内。MSI不仅在原位分析方面具有广泛适用性,而且在无需标记的情况下同时分析多个组分方面具有高通量和高效率的特点,这是其他原位分析技术所无法比拟的。近年来,基于MSI的优势,植物学家一直致力于开发一种完整而高效的空间多组学研究模型,以探索植物在环境中动态表型与稳定基因型之间的调控。这一空间多组学模型还被应用于确定植物多种生理和生化状态的微观代谢密码。植物中的代谢途径可以在组织、细胞甚至亚细胞水平上还原为三维结构。
近年来,基于MSI引领的空间多组学研究模型揭示了植物在不同应激和短期或长期应对机制下的代谢调整,这些机制反映了植物与不同相关微生物之间的代谢通讯差异,以及植物对不同类型外源物质的代谢途径差异。我们使用关键词「质谱成像和植物成像」在Web of Science和PubMed数据库中进行检索,并在2019年至今检索到60篇相关研究,本综述介绍了MSI技术的基本原理和实验程序,并总结了过去3年基于MSI的空间代谢组学在研究代谢物、植物生理和生物化学关系方面的主要应用,包括植物天然产物的空间生物合成途径、植物根与外部环境之间的物质交换、植物在非生物胁迫下的生理抵抗、植物在生物胁迫下的化学防御以及植物与微生物之间的相互作用。我们还从关键指标的角度讨论影响图像质量的因素,并展望了基于MSI引领的空间多组学研究模型结合遥感等现代表型技术,在未来可能取得的对植物在环境中动态表型与稳定基因型之间调控的突破。
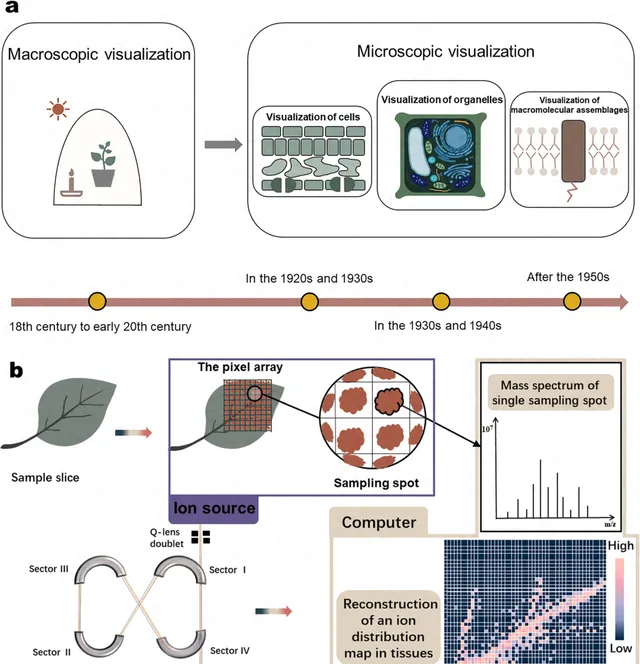
图1 基于MSI的植物微观可视化和植物代谢物空间表征的历史。(a)近100年植物微观可视化的发展(可视化从宏观到细胞、到亚细胞、到大分子的发育过程);(b)MSI重建代谢物分布图的流程。
2 质谱成像技术的原理、实验步骤和影响因素
2.1 MSI 原理和不同的电离类型
质谱成像仪主要由离子源、质谱分析器、探测器和数据处理系统组成。相较于其他质谱仪,质谱成像仪可利用特殊的离子源记录待测离子的空间坐标,并通过计算机的存储、识别、筛选和分类功能,重建离子在组织中的分布图。除了常规参数如采集速度和质量分辨率外,还考虑了空间分辨率和成像的可重复性。MALDI、DESI和二次离子质谱(secondary ion mass spectrometry,SIMS)是分析植物小分子组分的三种最常用的离子源。表1提供了几种离子源的具体参数。
表1三种常用离子源的比较

2.1.1 基质辅助激光解吸/电离 (MALDI)
由于离子源的电离过程,MALDI-MSI所使用的样品在MALDI-MSI中有一个额外的步骤,即在样品上喷涂基质。但总体方法相似。典型的MALDI-MSI工作流程如图2b所示。通过适当的方法获得的适当厚度的组织切片被放置在铟锡氧化物(ITO)涂层的载玻片上。选择适当的MALDI基质,并均匀喷洒或升华到组织表面,组织中的分析物最初在溶剂中溶解。随着溶剂的蒸发,基质和分析物根据分子间力(包括氢键、卤键、范德华力和π-π堆积)形成超分子结构的晶体。带有共结晶样品的载玻片被放入真空干燥器中,以防止吸湿。然后,带有共结晶样品的载玻片被放入MALDI质谱仪进行分析。MALDI-MSI的电离过程发生在高真空环境中。首先形成基质离子,然后在激光束照射在玻片圆形区域上的情况下,吸收能量的分析物被汽化。在气化过程中,代谢物通过基质离子与分析物之间的碰撞被离子化并进入四极杆分析器。通过扫描整个样品,激光束可以跨越整个组织切片进行原位分析。在电离过程中,激光脱附具有强烈的电离效应,但来自组织的分析物可以通过基质离子的间接电离。相对较温和的电离保持了电离效率,而空间分辨率主要取决于激光斑的大小,通常为5-10μm。大气压-基质辅助激光解吸/电离(AP-MALDI)是一种类似于MALDI的电离源;两者都具有相同的样品预处理步骤,并且都需要喷洒基质来协助分析物的电离。AP-MALDI与MALDI的区别在于,分析物的汽化和电离发生在大气压下。AP-MALDI是一种较为柔和的电离方法,更适用于不稳定化学成分,并且可以产生更稳定的离子流。AP-MALDI的大气压电离使基质离子能够与分析物离子形成更均匀的电子云,并且空气对流起到冷却效果,降低了由激光灼热引起的分解损失。此外,AP-MALDI的灵敏度主要由大气压电离源和真空质谱分析器之间的离子传输效率决定,这通常需要选择具有更大喷嘴和更高样品消耗的AP-MALDI。
2.1.2 解吸电喷雾电离 (DESI)
DESI是一种电离源,其原理在理论上与电喷雾电离(ESI)相似(图2a)。通过使用高电压电场对溶剂进行带电。使用喷雾气体(N2)的气动雾化器不断向组织中喷洒溶剂液滴,导致其路径中的电荷积累和溶剂蒸发。这导致了重复的库仑爆炸和更小的带电溶剂液滴。带电的小溶剂液滴通过快速移动的雾化气体(N2)引起的气体喷射影响组织表面。带电液滴在样品表面的撞击点溶解代谢物,然后碰撞、分解并喷射进入喷嘴口。在这些过程中,撞击点处的分析物被电离,而溶剂继续挥发,空间分辨率约为50μm。MSI的定位精度和可重复性受到溶剂毛细管的几何特征(如刚度和尖端质量)的显著影响。此外,通过优化溶剂组成、流速和几何因素,还可以在不改变喷雾器结构的情况下在一定程度上提高DESI的空间分辨率和性能。
2.1.2 二次离子质谱 (SIMS)
SIMS中的电离是在组织表面发生的阴离子级联碰撞过程(图2c)。在高真空中,离子发射器将一束具有高动能的离子直接照射到样品表面。在样品表面的撞击点上,被称为电荷初级碰撞离子的表面粒子在电离过程中获得动能,并发生异位碰撞产生二次碰撞离子。小的二次离子被溅射到注入器中,并在高速离子轰击过程中到达质谱分析器。纳米级二次离子质谱(Nano-SIMS)和飞行时间(TOF)-SIMS是两种常见的质谱技术。通过在样品表面进行高剂量离子轰击,nano-SIMS可以达到的最高横向空间分辨率为50纳米。使用较低的离子剂量进行电离过程的TOF-SIMS无法达到这种高空间分辨率,但可以将对样品表面的损伤降低到单个原子或分子层的1%。此外,TOF-SIMS适用于有机化合物和单原子离子的分析,因为既可以检测低质量片段,也可以检测相对较高质量的片段,显示了广泛的应用。TOF-SIMS还可以进行多组分-多系统的同时分析、深度剖析分析、3D成像等功能,使其在生物成像中得到更广泛的应用。
2.2 植物样品MSI的实验程序
在通用的二维质谱成像技术中,通过简单的预处理方法(如切片或压印),尽可能在植物样品中保留所有代谢物在同一平面上。载有该平面的载体被放入MSI仪器,以预设的空间分辨率进行数据采集。空间分辨率表示在同一平面上两个像素可以被区分的最小距离,直接反映了仪器对空间区分的能力。空间分辨率通常通过坐标方块矩阵进行可视化。每个坐标点中的代谢物可以通过不同的离子源使用不同的电离方法进行收集,并最终进入质谱分析器以收集相关的质谱信息。为了创建总离子流程图,将每个像素中发现的所有片段离子及其独特的电荷比率汇总。计算机软件可以提取在误差允许范围内具有相同m/z值的每个离子的所有像素位置,并将它们存储在每个离子坐标数据集中,用于重建离子分布图(图1b)。
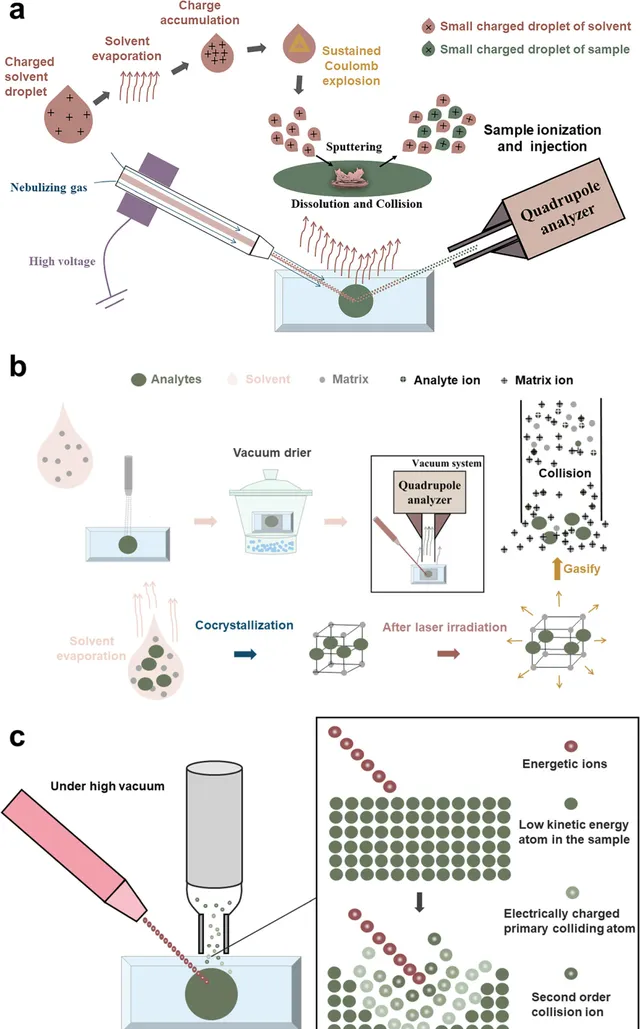
图2 基于MSI的植物微观可视化和植物代谢物空间表征的历史。(a)DESI原位电离示意图(样品表面分析物在溅射到进样口之前在带电微滴的作用下电离的过程);(b)MALDI的显微演示(MALDI操作程序和基质辅助电离原理); (c)基于离子级联碰撞的 SIMS(离子束轰击下样品表面分析物电离的微观过程 )。
2.3 质谱成像的影响因素
MSI可以提供化合物的传统质谱信息和离子的空间坐标信息。基于期望获取两种相应数据的前提,我们需要从质谱和空间两个维度的关键指标来理解影响MSI的因素。影响质谱成像图像质量的主要变量包括样品完整性、分辨率、灵敏度和成像重复性。
2.3.1 样品制备
对于许多使用冷冻微切片法作为预处理方法的样品,获取具有适当厚度、完整形态且无污染的切片是成像质量的关键因素。切割的组织需要在适当的刚度状态下准备,并且能够被冷冻微切片机轻松切割成微米级的切片。有时,从生长中的植物中切取新鲜样本,其含水量过多或组织不够紧实,容易被冷冻微切片机的刀片碾碎。这些新鲜组织可以被冷冻在-80°C以稍微降低水分含量,并增加植物组织的紧密度,或者它们可以嵌入到适当的物质中。目前,最优切割温度(optimal cutting temperature,OCT)化合物是最常见的嵌入介质之一。然而,如果组织完全嵌入到OCT中,它将成为导致离子抑制问题的污染物,影响质谱分析。一些最近发现的嵌入介质可以在不影响质谱数据的情况下协助冷冻切片,包括羧甲基纤维素(carboxymethyl cellulose,CMC),这是纤维素和明胶、明胶、冰、蛋清或煮沸介质的混合物,提供了解决冷冻切片问题的潜力。此外,一些木材组织由于含水量低、纤维含量高、无粘性以及难以获取新鲜样本,过于坚硬难以切割成切片,需要新的技术或新的包埋介质。
2.3.2 空间分辨率
MSI中最关键的一个方面是其空间分辨率,这是理解MSI数据的一个重要概念。对于2D成像,需要考虑横向空间分辨率;这个分辨率表示在同一平面上两个像素可以被区分的最小距离,直接反映了仪器对空间区分的能力。仪器的空间分辨率是基于一般认识的,即样品区域和两个采样点之间的最小距离都不会影响计算机能够识别的坐标。通常报告的预设分辨率实际上是一个2D像素数组,它实际上是样品表面上采样点的密度。MSI仪器在每个坐标点获取并记录提取的分析物离子的质谱,包括提取分析物离子的所有坐标点的位置,以便可以恢复原始组织中目标离子的分布(图1b)。
基于样品斑点大小和间隔距离这两个主要因素,有许多技术用于提高MSI的空间分辨率。根据过去10年的研究,使用MSI离子源结合小孔、多焦距透镜、光束扩展器或更改光束传输选项,已经可以定位个别细胞或亚细胞结构,甚至在三维中实现。这有可能取代费力的连续切片。使用自动聚焦光谱对不规则叶子样本进行逐层采样,创建亚微米深度空间分辨率,从而实现 3D MSI。根据最近文献的检索,3D质谱成像在植物生理学中并没有广泛应用。
这有潜力替代繁琐的连续切片。Dreisbach等使用自动对焦光谱学逐层对不规则叶片样本进行采样,创建了亚微米深度的空间分辨率,从而实现了3D MSI。根据对近期文献的搜索,3D质谱成像在植物生理学中并没有广泛应用。
2.3.3 质量分辨率和质量灵敏度
质谱仪在两个离子之间能够分辨的最小m/z差异反映了仪器的质量分辨率;这个分辨率是影响成像离子通量和质量准确性的最关键因素。相等大小的两个峰被认为在半峰的高度处分开,而在半峰的高度处的峰宽是质量分辨率的数学表示。质量分辨率的应用意义主要体现在质谱仪对不同m/z离子的分辨能力、识别离子的m/z范围的大小以及待测离子的质量准确性。这些对于不同类型的质谱成像分析仪器的质量至关重要。以下讨论了几种常见的质量分析仪器,表2提供了几种质谱分析仪器的具体参数。
表2 不同质量分析仪的性能(质量分辨能力和质量精度主要针对m/z>1000)
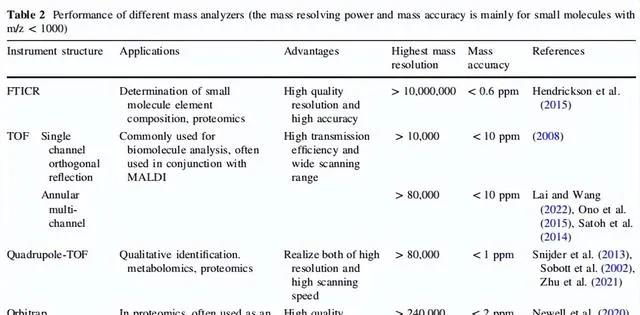
在相同的初始位置经过脉冲直流电场后,待测离子具有相同的初始速度并在飞行管中自由移动;然而,由于其不同的m/z值,离子到达探测器的时间是不同的,基于此概念TOF不断得到改进。由于正交速度可以使待测离子在垂直于原始行进速度的方向上具有相同的初始速度。在离子在第一个垂直方向的初始速度降为零后,由于另一个电场,离子的速度调整到相反的方向,然后离子在第二个飞行管中自由移动以到达探测器。这个过程基本上使离子的飞行长度增加了一倍,而不会增加仪器的尺寸。在以上原理的支持下,单通道正交反射飞行时间质谱仪的质量分辨能力可以超过10000。随着时间的推移,由于环形多通道TOF-质谱仪的出现,这种类型的质谱仪的质量分辨能力已经提高到了超过80000;然而,由于通道的延伸,m/z检测范围也相应减少了。目前,TOF经常与自身或其他质量分析仪器串联以实现更高的空间分辨率、更宽的m/z检测范围、更准确的质量检测准确度和更灵活的工作模式。
Orbitrap也是一种高分辨率质谱仪。这种质谱仪灵感来自Kingdon陷阱并可视为其升级版本。在20世纪90年代,这种基于静电场的轨迹陷阱被认为对于质谱分析是有用的。两个轴对称电极嵌套在一起形成一个空间电场,外部电极为正,中心电极为负。通过精确设计这两个嵌套电极的形状,待测离子在电场的作用下以垂直于电场线的角度进入,离子同时进行稳定的轴向旋转和径向振荡。因此,围绕中心电极形成了稳定的离子轨道。精确的离子m/z可以从与初始离子速度和位置无关的轴向旋转频率中得出。基于这个Orbitrap,可以实现高质量分辨率。目前,Orbitrap在m/z 200处可以提供高质量分辨率(>240000),质量准确度<2 ppm;然而,其相对较慢的数据采集速度是一个限制。OrbiSIMS是一种新型质谱仪,它结合了TOF的快速采集速度和Orbitrap的高质量分辨率,突破了两者的限制。
目前,傅里叶变换离子回旋共振(Fourier transform ion cyclotron resonance,FTICR)质谱仪提供了最高质量分辨率。这基于运动点电荷在均匀电磁场中的电磁场理论。待测离子在均匀磁场中加速其回旋运动,通过时变电场的作用被激发到更大的轨道,然后被检测到。基于质谱仪原理的紧密拟合,通过共振修正了离子回旋运动的轴线,产生了可检测的电信号的空间干涉。旋转的离子在极窄的旋转半径内可以获得非常大的平移能量,这是FTICR超高质量分辨率的关键。
总之,离子采集效率、空间分辨率、成像重复性、质量分辨率和质量灵敏度等参数需要被用作关键影响指标。在大多数情况下,速度和灵敏度不能同时达到最佳状态;因此,需要根据实验需求选择速度或准确性,另一个则需要被牺牲。样品采集速度主要由像素点面积、像素点之间的距离和每个像素点的采集时间决定。通过减少像素的数量(增加像素点的面积或增加它们之间的距离)或减少采集时间,可以获得更高的采集速度。前者将导致较低的空间分辨率,而后者将要求质谱仪具有更高的灵敏度。对微代谢物的高灵敏检测主要与离子源的电离效率、离子传输效率和质谱仪的离子检测有关。质量分辨率主要与离子源和质量分析器中的精密结构有关,影响电离/解吸和质量分辨率能力。
3 MSI在植物分子生理学中的应用
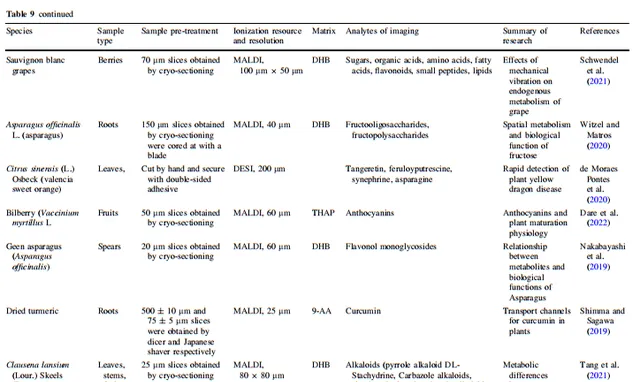
高等植物的基因组、转录组、蛋白质组和代谢组构成了弹性调控网络,负责维持它们的正常生命过程。除了利用代谢物的响应位置来确定生物合成途径中前体化合物、中间产物和产物之间的关系外,植物质谱成像(MSI)的空间代谢信息还能反映植物在内源和外源化合物方面的实际运行状态。通过分析植物组织中代谢物的聚集趋势,可以识别与其相关调控途径中连接的功能性物质。在本节中,我们详细总结了MSI在分析植物在非生物和生物胁迫下的胁迫耐受性和化学防御、植物与外界物质交换、植物与微生物相互作用、植物内源代谢物生物合成途径与生物学功能之间的关系、植物与微生物相互作用、植物杀虫剂代谢以及空间生理信息等应用方面的质量。所有植物质谱成像应用示例中涉及的离子源、空间分辨率、样品处理方法和成像分析物类型在表3、4、5、6、7、8和9中都有详细总结。
表3 植物在非生物胁迫下的耐逆生理
表4 生物胁迫下植物的化学防御
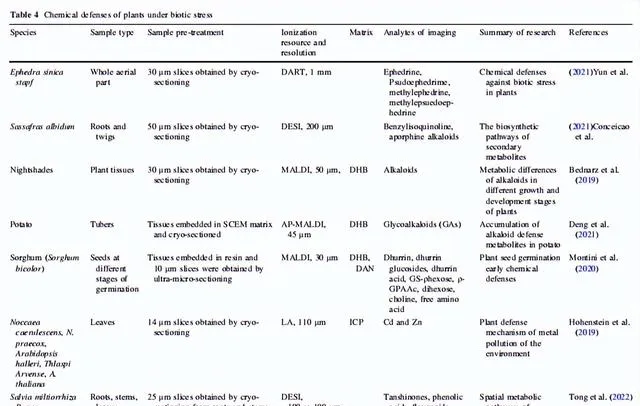
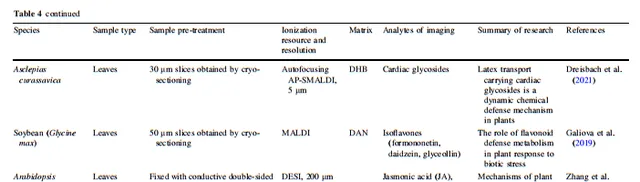
表5 植物根系与土壤的物质交换
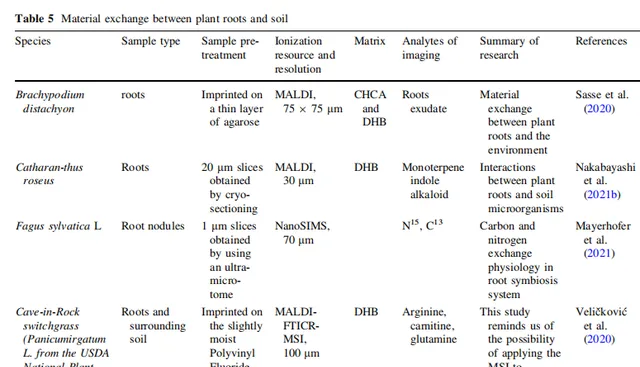
表6 植物-微生物相互作用
表7 植物次生代谢物的空间生物合成途径
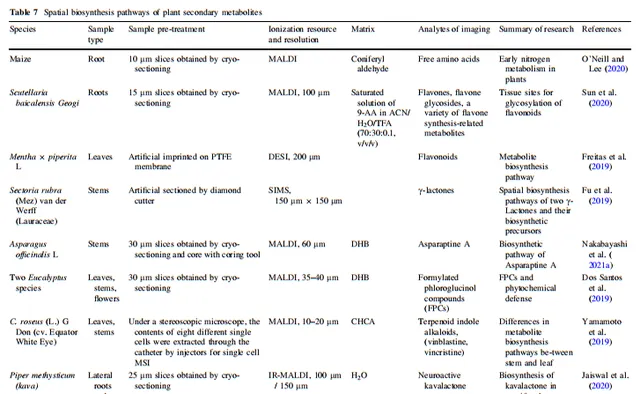

表8 外源物质在植物体内的代谢
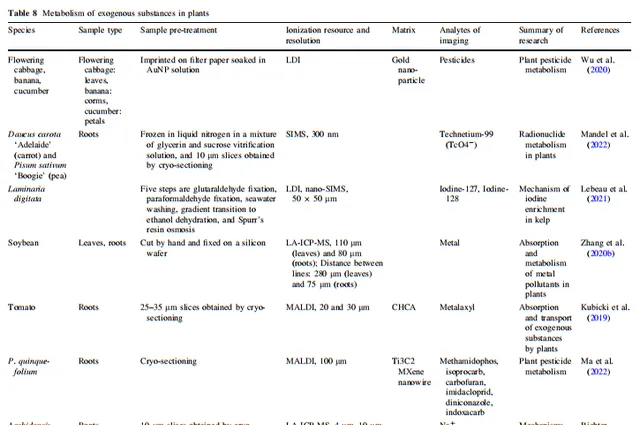
表9 其他应用
3.1 非生物胁迫下植物的胁迫耐受生理学
植物经历了适者生存的自然选择,呈现出一种区域性的生存趋势。植物区域性生长的进化筛选考虑到一些非生物因素,如温度、湿度、土壤盐度、酸碱度以及金属污染。当这些非生物胁迫因素刺激时,植物微系统中发生的一些最重要的适应性反应会增强植物的胁迫耐受性,并被认为是植物代谢调控的一部分。植物中的这些多重化学防御反应主要用于修复植物细胞的损伤,恢复植物细胞的稳态,并改变植物适应不同生长环境的方式。通过MSI在这些胁迫响应中重要的防御代谢物的组织分布中,部分揭示了植物抵抗力的生理机制。
在干旱、高盐、强光或升高温度的应对过程中,植物可以增加脱落酸的合成来促使叶片脱落并减少蒸腾。然而,植物细胞的水势由溶质势、压力势和衬质组成,是理解植物抵抗干旱机制的重要概念。面对各种非生物胁迫(如干旱、过盐和重金属污染土壤),植物通过增加小代谢物的生物合成,降低细胞的溶质势并维持渗透压,如在棉籽中观察到的那样。此外,在极端盐碱或干旱条件下,棉籽还通过增强甘油磷脂的生物合成来维持和补充细胞膜。在含有高汞和金含量的土壤中生长的木薯也出现了类似的情况;木薯在根表皮中具有相对较高的脂质含量。这些结果得到了Villette等人的研究的支持,该研究发现工业化合物及其代谢物主要被阻止在黑杨叶片靠近表皮的周围组织中。此外,Gupta等人提出,植物细胞中黄酮类化合物的积累影响了植物渗透压,并可作为耐盐大麦品种中的代谢生物标志物。
3.2 生物胁迫下植物的化学防御
植物必须应对包括病原体侵蚀和动物咬食在内的生物压力,以在生物圈中茁壮生长。在漫长的进化过程中,植物发展了多种次生代谢途径,合成和积累一些独特的防御化学成分,用于组织中抵御生物攻击。植物学家利用MSI收集生物碱、黄酮类化合物等与防御有关的化合物的空间代谢数据;然后利用这些数据解释植物在生物压力下的化学防御。
碱类化合物在许多植物中是非常重要的防御代谢物。这些碱类化合物储存在特定的植物细胞器中,受到胁迫信号的刺激后被释放并运送到作用位点,以抵抗真菌、细菌和其他病原体,以及昆虫捕食者。已经证明,尼古丁、α-石竹烯和α-茄碱等碱类化合物对多种啃食性和吸食性昆虫具有抵抗作用。士林和马钱子碱是两个碱类化合物的例子,它们与捕食者的酶结合,引起中毒或头晕。根据Yun等人的MSI研究,麻黄的侧枝中分布更多的麻黄碱和伪麻黄碱,这是因为前者更容易受到动物咬食。在檫木的地下部分,碱类化合物主要检测到在根的外表皮中,用于抵御草食动物、真菌、细菌等。然而,对于活体植物,特定防御代谢物的积累可能在生长和发育的不同阶段并不一致。Bednarz等人发现龙葵属植物的主要碱类在不同的生长和发育阶段发生了变化。随着储存时间的延长,马铃薯中四种有毒的配糖生物碱的合成增加,而在芽和果皮中的积累大于块茎组织,这可能是马铃薯安全发芽的一种防御机制。Montini等人发现dhurrin是一种重要的植物化学防御化合物,其生物合成和内源代谢发生在种子早期发芽阶段,以防止外界压力。
对黄酮类化合物的空间代谢分析是研究植物化学防御机制的另一个研究热点。Sun等人发现,黄芩根部的外周韧皮部是黄酮类化合物合成和糖基化储存的主要场所,在受到生物攻击时能够启动快速的防御反应,是其抵抗生物胁迫的重要生理表现。大豆和黄芩的防御机制非常相似,都是在叶薄壁组织中积累类黄酮来抵抗蚜虫。Tong等人发现 C-3 位点的羟基化激活了胁迫下植物中类黄酮的重新分配。
除了积累防御代谢物以防止损伤之外,植物还在受伤后经历代谢调节变化,以抵抗捕食者或恢复稳态。Dreisbach等人揭示了当马利筋叶片被动物咬食时,它通过分配和凝固橡胶乳液携带强心苷,以抵抗捕食者。植物激素也参与了植物受伤后的调节。Zhang等人发现茉莉酸(JA)、水杨酸(SA)、吲哚-3-乙酸(IAA)和脱落酸(ABA)都参与了植物叶片对机械应激的调节,其中JA的响应最高,而ABA没有明显的响应。SA的积累与JA及其前体化合物有关。
3.3 植物根系与土壤的物质交换
植物通过化学分子的语言与自然界进行交流,具体而言,植物主要通过与周围空气和土壤中的化学物质交换,与周围环境不断进行交流。土壤含有水分、许多元素和养分、丰富的微量元素,以及植物生长和发育所需的多样有机物。植物与其环境之间的物质长期交换也是整个植物生命周期中植物分子生理学的主要组成部分。近年来,许多植物的物质交换过程已经通过MSI进行了确定。
植物的地下部分(主要是根系和可能的地下茎)是植物与土壤相互作用的主要场所。由于根尖缺乏凯氏带和次生细胞壁,因此根尖是与周围环境进行化学交换的主要场所;这是通过MSI的应用所推断的,其空间分布结果显示根分泌物在根尖最丰富,而在成熟和伸长区域则分布较少。此外,根分泌物在成熟和伸长区域的少量分布表明,也通过活跃的运输或其他特殊通道发生了少量物质交换。此外,Veličković等人开发了一种根-土壤横截面膜印法,以研究活跃的幼根与土壤之间的物质交换;然而,由于整体根体积的限制,他们的研究未发现常见的土壤有机化合物离子。他们检索了整个根的空间代谢信息,包括阿拉伯半乳糖、苹果酸和其他关键代谢化合物,旨在确定植物对胁迫的抵抗基质。空间代谢组学在展示植物根部化学方面的潜力仍有待探索,主要是由于根-土壤横截面成像的困难。
3.4 植物-微生物相互作用
最近,利用基于MSI的空间代谢组成像技术,深入探讨了植物与微生物之间错综复杂的关系。不同的微生物群体以多种方式影响植物。Righetti等人使用AP-MALDI比较了福寿螺菌中各种代谢物的空间分布。他们还发现,亚油酸、α-亚麻酸及其相关代谢物在胚乳中积累,改变了其结构完整性。这一步是小麦福寿螺菌侵染的毒理学过程的一部分。然而,小麦中的许多防御机制也被激活,包括甘油三酯和半乳糖脂等信号分子;这些分子参与植物细胞膜修饰、植物激素受体激活、真菌识别和植物创伤后反应。此外,细胞壁开始积累更多的羟基肉桂酰胺,通过多糖交联与细胞壁成分结合,提高物理防御屏障。
基于植物对致病菌的适应性,许多研究人员通过MSI探讨了植物真菌的控制,为提高作物抗性提供了依据。例如,除了固氮作用之外,Hansen等人MALDI-MSI研究证实大豆结瘤还可以产生一些抗菌活性物质。Liu等人通过MALDI-MSI直接筛选了紫丁香中的抗菌活性物质,这为其他作物的真菌生物防治未提供证据。
MSI还为探索在一些植物中促进有益共生机制、促使微生物群落繁荣的机制提供了空间代谢信息。Brescia等人使用模拟根际环境的生长培养基,证明植物通过释放根际分泌物刺激Lysobacter capsici AZ78(AZ78)的细胞移动、生物膜形成和生物活性次生代谢产物的生物合成,从而影响土壤中的微生物群落,促进根系繁荣。对于促进生长的细菌,短柄草在根系中积累了类黄酮和酚酸等次生代谢产物,并增加了吲哚乙酸的合成以促进根系繁荣。由于MSI能够提供精细的空间代谢信息(植物与各种微生物以及微生物之间),因此在研究众多植物与微生物相互作用方面具有重要的潜力。
3.5 植物次生代谢产物
植物的糖代谢、脂质代谢、蛋白质代谢、核酸代谢、生物氧化和氧化磷酸化等生理过程相互关联,构成了维持植物正常生长和发育的基础。植物在应对环境或生物胁迫时,演化出许多复杂的次生代谢途径,形成了许多活性结构。植物的代谢网络已经得到广泛研究。许多类型天然产物的部分生物合成途径已经被确定,并发展成完整的化学结构衍生途径。MSI可以帮助将植物平面代谢网络与植物形态特征相关联,以理解植物的空间代谢特征。Yun等人检测到麻黄侧枝中麻黄碱和甲基麻黄碱等主要生物碱的分布高于主茎,而茎的内表面的分布高于外表面。这一结果可能与植物次生代谢产物对其自身的生物功能有关。O'Neill和Lee发现玉米根部的氮代谢在植物早期生长阶段显示出一定的亲本遗传性,通过获取玉米根部游离氨基酸的空间信息,了解在环境影响之前植物氮代谢的模式。研究人员旨在简化植物生长发育早期氮代谢(植物基本生理代谢)的研究,避免次生代谢的过度干扰。Sun等人证明了根韧皮是黄酮合成和储存的地方,也是黄酮苷酸的糖基化的地方。Li等人发现丹参根部的丹参酸在木质部合成,并部分储存在栓质和栓皮中。然后,丹参酸通过维管束穿过茎,并部分保留在髓腔中,最终从叶脉扩散到叶肉。Freitas等人通过DESI-MSI观察了一些在黄酮生物合成途径中关键化合物,这些化合物有潜力将黄酮生物合成途径与植物空间中的合成和代谢部位联系起来。Fu等人推测五种γ-内酯可能是红扇贝茎的射线薄壁细胞中同一生物合成途径的一部分;他们的研究展示了TOF–SIMS MS/MS成像辅助其他技术了解植物内源产物生物合成途径的潜力。Yamamoto等人通过代谢物的单细胞MSI发现了长春花的茎和叶之间的代谢产物的生物合成功能存在轻微差异。Revuru等人进一步提供了支持将skyrin作为金丝桃素前体化合物的证据,并通过MALDI-MSI假设后者的生物合成途径发生在金丝桃叶的黑色腺体中,而大黄酚对金丝桃素的生物合成不直接相关,因为它在叶中广泛分布。
3.6 植物体内外源物质的代谢
不同的植物可能对相似的外源物质拥有相似的代谢途径。具体而言,植物在漫长的进化历史中发展出代谢的多样性,但仍保留了一些相似的基础生理过程。例如,植物主要通过蒸腾作用吸收中性化学物质和非必需金属,因为植物吸收这些物质的量与蒸发的水量呈负相关关系。Kubicki等人利用MALDI-MSI证实了甲酯杀菌剂在番茄植物的常规吸水过程中是通过木质部和皮层完成系统传输的。同样,农药转运体从韧皮到木质部扩散,随着水在木质部中的运输,最终在叶缘积累。这一动态过程由Wu等人通过MALDI-MSI进行了深入观察。MSI分析还显示,许多金属污染物仍停留在韧皮之外。此外,尽管钠是植物的一种必需成分,但过多的钠也被阻止进入韧皮部。然而,植物的代谢也存在一些变异性。Mandel等人的SIMS单细胞MSI数据显示,植物对三种放射性核素的富集过程是不同的。锝(Tc)通过主动运输横向转移到木质部,并随着水一起运输,而铼(Re)被植物吸收较少,只在胡萝卜吸收后存储并在薄壁组织中运输。Lebeau等人通过DESI-MSI发现,在富含碘的海藻海带中,放射性核素I和常见同位素I具有不同的细胞外分布。这些外源代谢数据的获取为探索植物生理的外源调控提供了适当的基础信息。
3.7 其他应用
除了上述六大类应用之外,MSI还有一些其他的应用,这里进行了总结。Witzel和Matros使用MALDI技术探索草莓代谢物与其外观和口感之间的关系。De Moraes Pontes等人使用DESI-MSI寻找甜橙的病害标志物,以实现对黄龙病的快速检测。一些植物学家结合植物的空间代谢信息与其生物学功能,解释它们的代谢行为。综上所述,MSI 在探索植物生理和病理机制方面显示出显著的应用潜力。
4 MSI的局限性和未来前景
回顾近期文献,空间代谢组学能够系统全面地展示植物代谢物的空间分布,有效地将植物代谢与植物形态联系起来,从而为揭示植物分子生理学提供了丰富、多样和高通量的数据支持。尽管通过MSI实现的代谢物的分子定位在植物空间代谢研究中并非不可替代,但其高通量的定位能力结合代谢组学和样品处理的便利性在其他方法中是不可替代的。然而,不能忽视空间代谢组学的局限性。首先,并非所有植物组织都能够进行空间分析,因为从一些脆弱、干燥和坚硬组织中获取适当样本存在困难。其次,作为所有质谱技术的共同问题,离子抑制在MSI中也是一个重要考虑因素。因此,除了简单的切片处理外,一些研究人员还使用有机溶剂的样品预处理过程来去除杂质。然而,一旦预处理过程变得复杂,植物代谢物的变化就成为一个主要因素。第三,大量空间定位信息的获取是一个双刃剑,给研究人员在提高植物研究系统性的同时提取植物生理信息带来了困难。最后,植物生理过程的变化只能根据空间代谢组学提供的实际植物代谢物来进行推理、假设或验证。例如,许多植物代谢网络操作的动态问题无法通过空间代谢组学解决。
我们认为,MSI的这些局限性不会掩盖植物空间多组学研究模型在未来对植物生理特性、植物动态代谢、基因和可变表型之间的动态物质基础进行研究的潜力。表型以及转录组、蛋白质组和代谢组变化的准确识别和定位是植物获得性遗传研究的重要方面。通过多种高通量和高精度的组学方法,可以适当地表征植物的基因组、转录组、蛋白质组和代谢组。由于近距或远距传感工具的发展,现在可以精确分类植物表型。近距传感是指测量靠近植物的设备,如计算机断层扫描和磁共振成像,而远距传感则利用电磁波辐射、反射特性等方式远离植物进行非接触测量。MSI主导的空间多组学研究模型可以提供关于植物的跨组织和跨细胞代谢过程的信息,促进植物结构、生理学、形态等复杂表型与转录组、蛋白质组和代谢组的多样性之间关系的有效筛选。MSI主导的空间多组学研究模型预计将发展成为植物生理学解释和育种设计优化的有力工具,以及探索食用和药用植物农学和植物生理学的有力工具。











