编译:微科盟 梅子,编辑:微科盟 景行、江舜尧。
微科盟原创微文,欢迎转发转载,转载请注明来源于【转录组】公众号。
导读
土壤镉(Cd)污染日益严重,需要有效的修复策略。 本研究调查了外源茉莉酸(JA)对Cd胁迫下东南景天( Sedum alfredii Hance)的影响,旨在提高其植物提取修复效率。研究者首先评估了在Cd浓度为100、300和500μmol/L的环境中添加不同浓度JA的影响 。结果表明, 浓度为1μmol/L的JA最合适。这一浓度通过提高抗氧化酶的活性,有效地降低了ROS产物的水平 。此外,JA还能促进Cd的吸收和积累,同时明显改善植物的生物量和光合作用。然后, 研究者在300μmol/L Cd胁迫下使用1μmol/L JA处理东南景天,转录组分析揭示了一系列JA介导的在响应Cd胁迫中起重要作用的差异表达基因(DEGs)。这些差异表达基因不仅包括JA生物合成和信号传导途径,还包括编码影响抗氧化系统和光合作用功能的基因,以及与细胞壁合成、金属螯合和转运有关的基因 。 本项研究表明,JA处理能显著增强东南景天对Cd的耐受性和积累能力 ,为植物修复提供了一种前景广阔的策略,并加深了研究者对植物应对重金属胁迫的理解。
论文ID
原名: Jasmonic acid’s impact on Sedum alfredii growth and cadmium tolerance: A physiological and transcriptomic study
译名: 通过生理学和转录组学分析研究茉莉酸对东南景天生长和耐Cd能力的影响
期刊: Science of the Total Environment
IF: 9.8
发表时间: 2024年1月
通讯作者: 杨文浩
通讯作者单位: 福建农林大学资源与环境学院
DOI号 : 10.1016/j.scitotenv.2024.169939
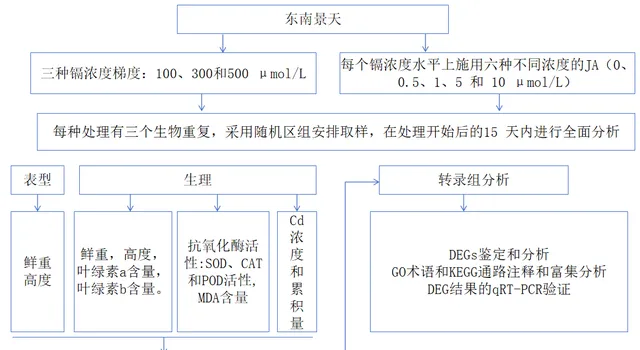
实验设计
结果
1 施用Cd和JA的植物生长情况
通过植物鲜重和高度的变化,可以清楚地看出JA对东南景天生长的影响。如图1(A和B)所示, 随着外源JA浓度的增加,东南景天的鲜重和高度都呈明显的上升趋势 。具体来说, 在施用10 umol/L JA的情况下,与施用0μmol/L JA的植株相比,不同Cd浓度下的东南景天鲜重增加了10.12%到29.44%。植株高度也有所增长,增幅从15.67%到7.13%不等 。值得注意的是, 东南景天在1μmol/L JA的条件下达到了生长的顶峰,鲜重达到2.66g,株高达到17.24 cm。
图1(C和D)显示了各种处理对东南景天光合色素的影响 。在所有探究的浓度中,Cd的影响主要是抑制作用,随着Cd浓度的增加,抑制作用逐渐减弱。 与0μmol/L JA处理相比,随着JA浓度的增加,不同Cd浓度下东南景天的叶绿素水平均呈现先升高后降低的模式 。 在1μmol/L JA浓度下,光合作用和色素沉着的增强效果最好, 尤其是在300μmol/L Cd浓度下,叶绿素a和叶绿素b的浓度分别达到0.27 mg/g和0.11 mg/g。
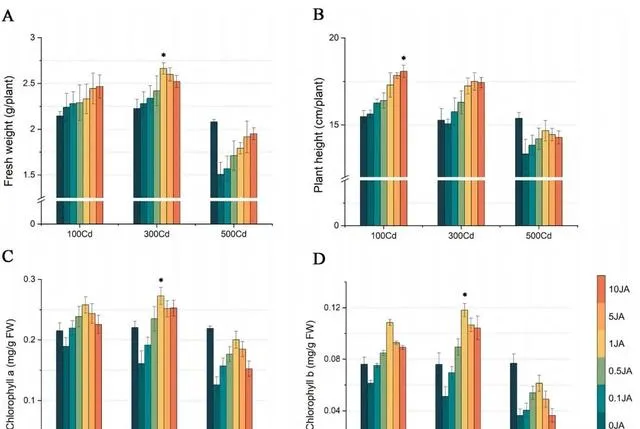
图1.Cd和JA处理对东南景天生物量和色素沉着的影响。 (A)东南景天的鲜重,(B)东南景天的高度,(C)东南景天的叶绿素a含量,(D)东南景天的叶绿素b含量。
2 施用JA对Cd胁迫下抗氧化酶活性和MDA含量的影响
研究者通过测量东南景天的MDA含量,仔细研究了重金属引起的氧化应激 (图2)。与CK相比,在不同的Cd剂量下,MDA含量上升了32.42%至65.16%。然而,加入较低浓度的JA(0.1、0.5和1 μmol/L)后,MDA含量大大减少。更确切地说,在不同的Cd浓度下,1μmol/L JA的用量比0μmol/L JA条件下分别减少了15.38%、16.38%和24.20%。相反,JA剂量越高(5μmol/L和10μmol/L),MDA含量越高,在Cd浓度为500μmol/L时,MDA含量明显增加,与1μmol/L JA相比,5μmol/L JA和10μmol/L JA处理的MDA含量分别增加了4.38%和12.21%。
同时, 研究者还分析了不同JA处理对东南景天中SOD、CAT和POD活性的影响,目的是减轻Cd诱导的胁迫 (图2)。结果证明,在所有测试方案中, 较低剂量的JA都能显著提高抗氧化酶的活性,在1μmol/L JA的情况下达到顶峰 。与不含JA的对应物相比,研究者注意到在各种Cd浓度下,SOD、CAT和POD活性分别提高了27.44%、25.48%和23.46%,38.82%、36.89%和30.61%,以及20.28%、22.90%和20.14%。然而,在5μmol/L JA和10μmol/L JA区间,抗氧化防御能力逐渐减弱,导致响应收到抑制。
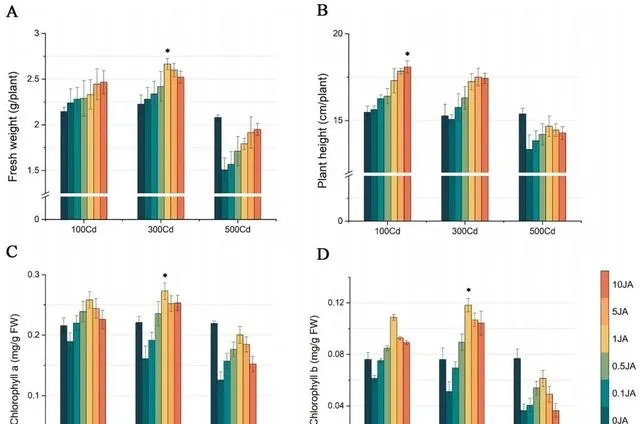
图2.Cd和JA处理下东南景天的MDA和抗氧化酶活性。
3 Cd胁迫下JA处理对Cd积累的影响
图3显示了东南景天显著的Cd积累能力,0μmol/L JA组的Cd含量分别为4803.7、7508.28和5447.92 mg/kg。引入JA能明显提高东南景天的Cd积累潜力。 研究者对暴露于100和500μM Cd条件下的东南景天进行实验,所获得的数据表明,随着JA含量的增加,东南景天中的Cd浓度也在持续增加。 在10μmol/L JA条件下,Cd浓度的峰值分别为6640.98mg/kg和5927.60mg/kg,在无JA条件下分别增加了27.68%和8%。在300μM Cd设置中,随着JA的增加,东南景天中的Cd含量开始上升,但随后出现下降, 在1μmol/L JA时达到顶峰,浓度为8639.34mg/kg (与0μmol/L JA相比上升了13%)。同时, 研究者对100μM和500μM Cd环境的东南景天研究表明,随着外源JA浓度的增加,东南景天的Cd积累量也在不断增加 。在10μmol/L JA条件下,100μM Cd和500μM Cd处理下的东南景天积累值分别为656.41μg/plant和290.42μg/plant,与0μmol/L JA相比,分别大幅增长了1.60倍和2.28倍。 300μM Cd+1μmol/L JA条件是东南景天对Cd积累最有效的条件,达到1009.72μg/plant的峰值(与0μmol/L JA相比提高了1.43倍) 。

图3.Cd和JA条件下东南景天的Cd浓度和累积量。
4 Cd胁迫下用JA处理的东南景天的转录组分析
为了进一步探讨Cd和JA处理下东南景天的生理和分子机制,研究者根据东南景天的总体Cd含量及其生理参数,选择300μM Cd+1μmol/L JA处理组进行转录组学分析。 转录质控数据见补充材料图S1和表S2,数据库注释数据见图S2。 研究者通过分析在JA和CK处理的地上部分中发现了7254个DEGs (图4)。其中, 3503个基因在地上部分上调,3751个基因下调 。相比之下, 根部的DEGs数量更为显著,共有10399个。其中绝大多数,特别是9554个基因被上调,只有845个基因被下调 。所有DEG都符合绝对log2(fold change)≥1和 P 值<0.05的标准。
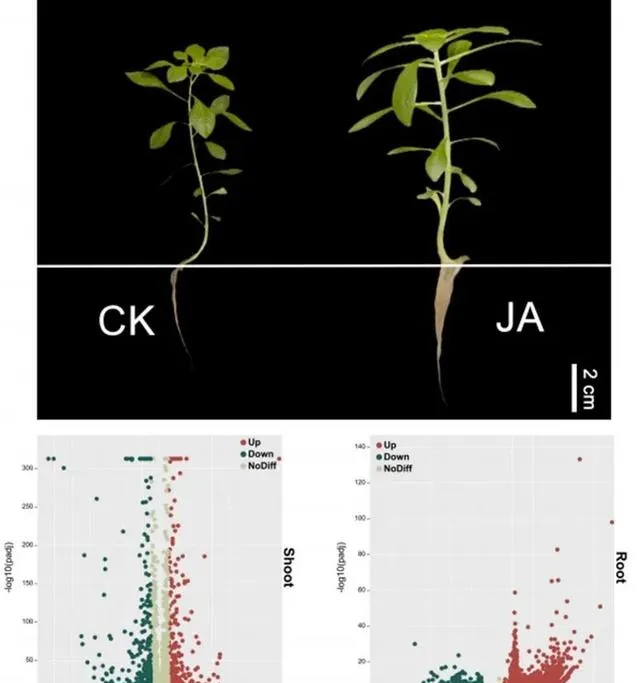
图4.东南景天在Cd和JA条件下的生长和差异基因的表达。
5 GO术语和KEGG通路注释和富集分析
在东南景天对Cd和JA处理的响应中,GO注释分析(图S3)揭示了DEGs在三个本体中的分布:生物过程(BP)、分子功能(MF)和细胞组分(CC)。 从TOP30 GO术语中,研究者可以观察到以下趋势:在用JA处理的东南景天的地上部分中,有23个GO术语被归入CC类别 , 主要与"细胞膜和细胞壁"基因有关。在BP和CC两类中,最富集的GO术语与"光合作用"和"金属和阳离子转运"有关 。而 在根分析中,在TOP30的GO术语中,有14个术语被归入生物过程(BP)类别,主要与氮化合物和胺的生物合成有关。13个术语被归入CC类,主要集中在与核糖体有关的结构上。CC类别还包括结构功能。
为了深入研究JA促进Cd胁迫下东南景天中DEGs的生物学特征,研究者利用KEGG通路数据库进行了通路富集分析 (图S1)。 结果表明,东南景天的KEGG代谢通路包括细胞组织结构和分子运输、信号转导、氨基酸代谢、脂肪酸及其衍生物代谢和光合作用等多个方面 。具体来说, 在地上部分,最主要的代谢途径是过氧物酶体、内吞作用、碳代谢和植物与病原体的相互作用。相比之下,根部的主要代谢途径包括裂殖体、RNA运输、碳代谢和脂肪酸代 谢。
6 DEG结果的qRT-PCR验证
为了证实转录组测序的准确性,研究者对来自芽和根的四个关键基因进行了qPCR检测,这些基因与多种Cd响应途径密不可分 (图S4)。所选基因主要与细胞壁合成、金属螯合和运输过程有关。它们的表达谱与Illumina测序结果非常吻合,进一步验证了DEG分析的可靠性。
讨论
1 JA在Cd胁迫下促进东南景天的生长
在本研究中,研究者观察到JA对东南景天在Cd胁迫下的光合作用有积极影响,表现为叶绿素-a和叶绿素-b的含量显著增加 (图1)。这与芽中的许多DEGs有关,包括参与光合系统I和II、光合电子传递链、F型ATP合酶和光合叶绿素-蛋白质复合体的DEGs(图5)。在光合作用过程中,光系统I和II被证明共同驱动电子传递链,协同完成光合电子传递。它们将光能转化为生长所需的能量和有机化合物,支持植物的生长和发育。然而,Cd胁迫会阻碍这一过程,降低光系统I的活性,增加ROS的产生,从而影响卡尔文循环和其他生物合成途径。 本研究发现,JA可通过调节PSII和PSI的基因表达,增强光合系统的稳定性和活性,这有助于植物更好地适应外部环境胁迫,保持健康的生长和发育 。 研究者还观察到,JA影响F型ATP酶多个亚基的表达 。这种酶是一种多亚基蛋白质复合物,通过质子动力催化ATP的合成或水解。 本研究表明,外源JA可以影响这种酶的各种亚基的表达,维持细胞内的pH平衡和能量供应,从而确保细胞的完整性 。这有助于植物在面临Cd胁迫时维持正常的细胞功能,从而促进其生长和适应性。

图5.Cd和JA条件下与东南景天光合作用相关的DEGs分析。
2 JA提高了东南景天对Cd胁迫的抗性
2.1 JA通过细胞壁提供额外的保护,降低Cd的毒性
Cd2+通过各种途径进入植物体内,对植物的生长和发育产生不利影响(图6)。植物细胞壁是最初的屏障,具有固定和捕获Cd2+的能力,从而减少其在细胞内的积累。 在植物的地上部分,研究者观察到参与木质素生物合成的CCR(DN81890_c3_g5和DN78139_c1_g1)表达上调,这可能会增强细胞壁强度并限制Cd2+的渗透 。此外,其他一些与木质素合成相关的实体,包括PAL(DN81699_c1_g5)和4CL(DN86000_c4_g4)的表达上升,也被证明有助于增加细胞壁密度,阻碍Cd2+的渗透。FAR(DN83112_c3_g1)和BGLH(DN80382_c1_g4)表达水平的升高可能会影响细胞壁的结构和稳定性。与地上部分相比, 根主要涉及与多糖和果胶合成有关的关键因子 ,如Cals(DN87352_c0_g1)和XTH(DN84935_c0_g1)。这些实体通过优化细胞壁的多糖结构,增强了对Cd的抗性。此外,PE可能通过调节细胞壁中果胶的合成来调节其稳定性和阻断Cd2+的能力。JA似乎可以通过上调CHI(DN83308_c1_g1)和EXP(DN145391_c0_g1)来增加细胞壁的可塑性和扩张性,以对抗Cd胁迫。 研究者还注意到,EXT(DN75005_c0_g2)家族的下调有可能导致细胞壁更具弹性和适应性,更适于承受外部压力,而不仅仅是增加硬度以抵御Cd的侵入 。同时,LAC(DN84135_c1_g2)和CAD(DN141324_c0_g1)参与了木质素合成的调控。CAD表达的增加意味着植物在应对Cd胁迫时强化细胞壁结构的策略,而LAC表达的减少则表明特定木质素生物合成途径可能受到限制。预计这种平衡可保持细胞壁的强度和柔韧性,而不会过度硬化。
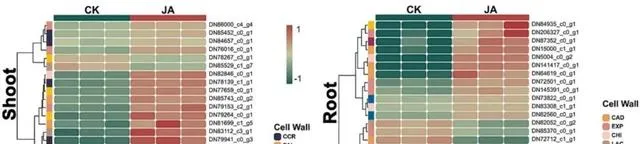
图6.在Cd和JA条件下参与细胞壁合成的DEGs。
总之, 研究者认为外源JA显著上调了一系列与细胞壁合成和修饰相关的实体,增强了植物对Cd胁迫的抵抗力,说明JA在细胞壁响应机制的调控中发挥着关键作用。 这些修饰将导致细胞壁更致密、更坚固,从而有效抑制Cd2+的渗透。
2.2 外源JA可通过抗氧化系统提高东南景天的抗逆性
植物进化出了非酶和酶的防御机制来抵御氧化应激,以应对具有挑战性的环境。先前的研究表明,植物激素JA能够刺激SOD、CAT和APX等抗氧化酶的合成,同时抑制ROS的产生,从而增强植物对Cd的耐受性。最新研究发现,在Cd胁迫下,JA合成基因AtAOS可刺激内源JA的产生,从而进一步抑制Cd引发的ROS生成,减轻Cd对植物生长的抑制作用,增强植物对Cd的抗性,本研究证实了这些发现(图7)。 研究者发现,添加外源JA能显著提高东南景天中参与抗氧化系统的几个关键基因(包括SOD、APX、GPX、PRX、CAT和GR)的表达水平,这说明JA在调节植物对Cd胁迫的响应,尤其是在建立和增强抗氧化防御系统方面发挥着核心作用 。总之,JA可通过多种途径增强东南景天的抗氧化防御机制,包括上调参与清除ROS的基因和促进建立更复杂的抗氧化网络。这有助于植物在Cd胁迫下保持稳定和有生命力的内部环境,提高其抗性。
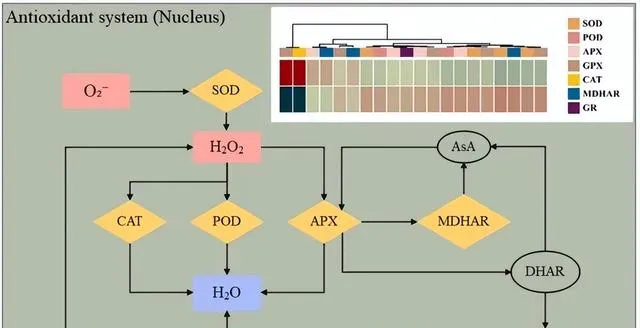
图7.在Cd和JA条件下参与抗氧化系统的DEGs。 O2-(超氧阴离子)、pO2(过氧化氢)、pO(水)、SOD(超氧化物歧化酶)、CAT(过氧化氢酶)、POD(过氧化物酶)、APX(抗坏血酸过氧化物酶)、GPX(谷胱甘肽过氧化物酶)、ASA(抗坏血酸)、MDHAR(单脱氢抗坏血酸还原酶)、DHAR(脱氢抗坏血酸还原酶)、GSH(谷胱甘肽)、GR(谷胱甘肽还原酶)。
3 JA通过调节金属螯合和转运蛋白促进东南景天对Cd的吸收和积累
通常情况下,当Cd2+穿过细胞壁进入细胞时,植物会启动一系列生化反应来降低Cd的毒性。最主要的反应是螯合,即利用小分子配体与Cd2+形成稳定的复合物。 在本研究中,研究者观察到外源JA显著调节了几个编码与金属螯合相关的蛋白质的基因的表达(图8),揭示了JA增强了东南景天螯合Cd2+的能力 。 研究者首先注意到PCs(DN77959_c1_g1)和MT(DN76429_c1_g3)表达的上调 。由PCS基因编码的蛋白植物螯合素能促进植物螯合素与Cd2+复合物的形成,有效降低细胞中游离Cd2+的浓度,从而减轻Cd的毒性。研究表明,当过表达来自苎麻( Boehmeria nivea )的新型植物螯合酶基因BnPCS1时,拟南芥对Cd2+的耐受、积累和转运能力都会增强。金属硫蛋白(MT)编码金属硫蛋白,有助于维持细胞中金属离子的平衡和解毒。虽然SaMT3主要与锌和铜形成复合物,但Zhao等人的最新研究发现,东南景天中的SaMT3具有Cd螯合能力。通过异源表达烟草金属硫蛋白基因NtMT2F,拟南芥对Cd胁迫的耐受性明显增强。此外, 研究者还观察到NAS(DN85360_c2_g5)、GST(DN82774_c0_g3)、Cys(DN148504_c0_g1)和OAS(DN218485_c0_g1)的协调上调。 值得注意的是, NAS通过烟碱胺的合成发挥了重要的金属螯合剂作用。它的上调表明,JA增强了金属螯合过程,从而减轻了地上部分的Cd毒性,并限制了根系对Cd2+的吸收 。另一方面,Cys和OAS是编码参与半胱氨酸合成(GSH的前体)功能的关键基因。因此,这些基因的上调很可能是放大GSH合成途径的关键,从而有效地管理Cd的封存和储存。
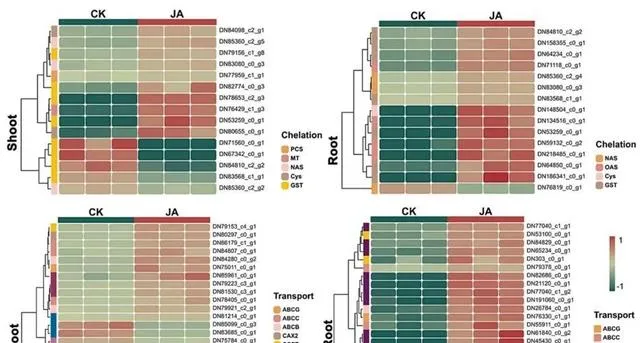
图8.Cd和JA条件下参与金属螯合和运输的DEGs。
金属转运蛋白在高积累植物吸收和转运金属的过程中起着至关重要的作用。尽管Cd不是植物的必需元素,而且在植物系统中也没有建立专门的转运途径,但转录组数据显示,外源JA能够通过增强其他二价离子通道(如Zn、Ca或Fe通道)中Cd2+的转运能力来调节东南景天的Cd积累。 为了系统分析这些基因在Cd管理中的作用,研究者将它们分为三大功能类别:(1)液泡中的Cd储存;(2)木质部和细胞质之间的Cd相互运输;(3)从根到芽的Cd长距离运输。
液泡是植物细胞中Cd2+的主要储存场所,其中VTL在Cd2+从细胞质转移到液泡的过程中发挥着关键作用。VTL是一种关键的铁转运蛋白,控制着Fe2+在细胞质和液泡之间的转运。有趣的是,虽然OsVTL1和OsVTL2已被证明参与了Fe2+的运输,但它们的过表达导致Cd胁迫下Cd在液泡中大量积累,这表明它们可能有助于螯合Cd和维持植物体内的金属平衡。 本研究结果表明,JA可选择性地调节VTL家族基因的表达,特别是通过上调VTL1(DN68490_c0_g1)在地上部分的表达,而降低VTL1(DN68490_c0_g1和DN81816_c1_g1)在根部的表达 。这种差异表达可能揭示了JA在调节Cd反应中的多层作用。例如,地上部分VTL的上调可能会促进更多的Cd被螯合到液泡中,从而增强植物对Cd胁迫的耐受性,从而降低Cd的毒性。同时, 根系中VTL1的下调可能是一种负反馈机制,旨在减少Cd的吸收和/或积累,从而保护根系免受Cd胁迫的损害。
在东南景天中,细胞质和木质部之间的Cd双向运输在调节Cd在植物各部分的分布中发挥着重要作用。CAX、NRAMP和HMA蛋白家族在这一过程中发挥了核心作用,确保了Cd的高效和平衡流动。其中,CAX是一种钙离子转运蛋白,对维持细胞内Ca2+的平衡和调节植物体内Cd2+的转运有着至关重要的影响。Zhang等人在东南景天中发现的SaCAX2h蛋白已被证明可调节Cd从细胞质到木质部的流动。值得注意的是,CAX的表达在地上部分和根部都受到外源JA的调控,这意味着 JA有助于提高东南景天对Cd的转运能力 。NRAMP基因家族被广泛认为能介导Cd2+和Fe2+的转运。 根据研究者的观察,NRAMP2和NRAMP5在Cd在细胞质和木质部之间的双向转运过程中表现出显著的表达差异。 具体来说,N RAMP5限制了Cd从细胞质向木质部的移动,而NRAMP2则促进了这一过程 ,这与Tian等人和Ge等人的研究结果一致。虽然HMA蛋白家族最初主要与Cu2+和Zn2+的转运有关,但也有研究表明它也参与了Cd2+的转运。特别值得注意的是, 研究者观察到HMA5基因在地上部分和根部都出现了上调。这可能表明外源JA通过调节HMA5影响了Cd在细胞质和木质部之间的转运,进一步优化了Cd在植物体内的分布 。
除了Cd在液泡和细胞质中的运输外,Cd通过木质部从根部向地上部分的移动也是一个至关重要的过程,这一过程确保了Cd在植物不同部位的有效分布,而YSL和ABC转运家族在促进这一过程中发挥了关键作用。黄色条纹样(YSL)转运体属于寡肽转运体(OPT)家族,主要负责在植物细胞膜上转运金属-植物苷元。研究表明,YSL家族参与了Cd的吸收及其从根到芽的长距离运输。 在本研究中,研究者观察到YSL12(DN82418_c1_g2)基因表达上调,这表明该基因在外源JA的影响下可能更加活跃,从而增强了其在金属-植物苷元转运中的作用 。这可能会增加Cd-植物苷元的吸收,进一步影响Cd2+的整体运输和分布。同时,YSL6(DN75784_c0_g1)表达的下调可能表明其在这一过程中的功能受到了限制。考虑到已知YSL6参与了Cd2+的长距离转运,其表达的降低可能为这一Cd2+转运途径提供了一种特有的保护机制。此外,ABC转运体蛋白家族也在木质部内转运Cd2+的过程中发挥作用。在这个家族中,ABCB、ABCC和ABCG亚家族被认为最直接参与Cd-植物苷元的转运。AtABCC1和AtABCC2在调节Cd从根到芽的转移过程中起着至关重要的作用。同样,OsABCG36和OsABCG43的高表达与水稻木质部内的Cd迁移有关。Naaz等人在大豆中发现,ABCB、ABCC和ABCG运输蛋白能够调节Cd2+的吸收和分布,帮助维持植物体内的Cd平衡。这与本研究结果不谋而合,因为 研究者检测到,在外源JA的作用下,东南景天的地上部分和根部ABCB、ABCC和ABCG的表达都出现了上调,这证实了ABC转运蛋白家族在通过木质部将Cd从根部转运到嫩枝的过程中发挥了关键作用。
此外, 研究者还观察到东南景天芽中COPT和Nipa的表达发生了变化 。这些是Cu2+和Mg2+的转运体,大量研究表明,Cd2+通过模仿Cu和Mg离子的化学性质,从而"欺骗"这些转运体,通过这些通道渗入植物细胞。 在本研究中,JA的引入明显增加了这些通道的表达量,促进了Cd2+向细胞内的更大迁移。
总之, JA在控制东南景天中的Cd2+转运方面发挥着明确的核心调节作用 (图9)。这种作用超出了对一个或少数几个分子通道的影响, 它协调了跨膜转运蛋白家族的广泛范围,并突出了其在植物应对Cd胁迫过程中的多方面功能 。最初, JA通过引导几个家族(包括CAX、VTL、COPT、Nipa和HMA)的表达动态,引导Cd2+在细胞内外的移动和储存 。例如,它通过促进金属离子向液泡的转运来支持维持细胞内离子平衡。这种作用既能保持植物细胞结构的完整性,又能减轻Cd过度积累的毒性影响。 随后,JA通过增强NRAMP和YSL家族选择性基因的活性,控制植物根部对Cd的吸收以及Cd在根部和叶片之间的长距离转运,从而在Cd诱导的胁迫下保护植物的生理结构和生长潜力 。 最后,JA还能刺激ABC家族的特定元素,加强植物的Cd排斥机制。 这些基本功能在Cd的吸收和清除中起着关键作用,确保了细胞内外物质的无缝运输,从而保护植物免受Cd胁迫的有害影响。
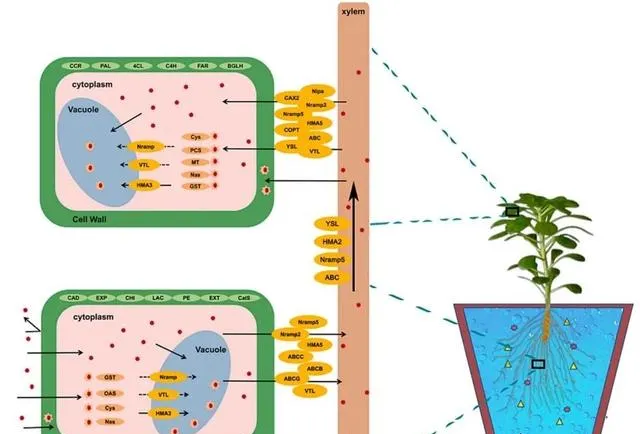
图9.在Cd和JA条件下参与Cd2+的DEGs示意图。
东南景天对Cd的吸收和积累受外源JA的显著影响。在Cd的吸收过程中,当Cd2+进入细胞质时,会与GST、NAS、CYS和PCS等蛋白质形成稳定的螯合物,提供主要的吸收通道。随后,CAX家族在Cd的细胞内积累过程中发挥了重要作用,促进了Cd从细胞质向液泡的移动 。 同时,NRAMP和HMA家族为Cd在细胞质和木质部之间的循环提供了核心途径,进一步加强了Cd在植物体内的积累 。 在木质部,Cd的向上运输主要由YSL和ABC家族成员完成 。值得注意的是,在这些关键过程中, HMA和NRAMP家族都受到JA的调控,这突出表明了JA在整个Cd吸收和积累机制中的决定性作用。
4 JA生物合成和信号传导在东南景天的生长和防御中的关键作用
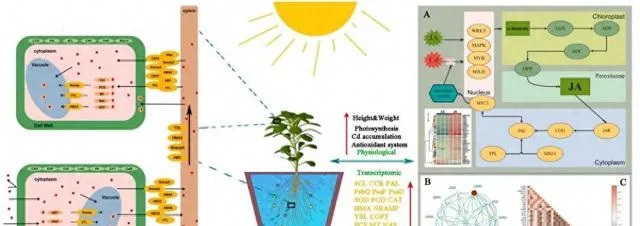
在Cd胁迫条件下,植物体内JA的合成和信号转导对植物的生长、抗逆性以及Cd的吸收和积累至关重要。当植物面临Cd胁迫时,它们的最初反应是产生大量活性氧(ROS)。ROS可被视为一种细胞警报,表明细胞内出现了胁迫。为了有效应对这种情况,植物会激活各种信号机制。在这一过程中,WRKY、bHLH、MAPK和MYB等转录因子起着关键作用(图10A)。特别是在有外源JA的情况下,这些转录因子的活性会增强。例如,已知WRKY46可调控JA相关基因对Cd胁迫的反应,MAPK信号通路被激活后可进一步调控JA相关基因。 本研究表明,Cd胁迫下转录因子WRKY46、MAPK3和MYB的表达上调,而WRKY4、WRKY50和bHLp5的表达下调。这种变化强调了外源JA对特定信号通路的调控影响,可能会增强东南景天内源JA的生物合成 。
由Cd诱导的转录因子在外源JA的存在下表现出对JA生物合成的积极调控作用 (图10A和B)。具体而言,植物体内LOX、AOS、AOC和OPR3基因的表达量显著增加,表明外源JA对JA生物合成有促进作用。 JA及其衍生物(如JA-Ile)在生物合成后可作为信号分子,启动特定的转录因子。这些转录因子进一步调节与JA反应相关的基因的表达,从而影响抗逆相关蛋白的合成 。 当JA-Ile与COI1结合时,JAZ蛋白降解,释放并激活MYC2转录因子。随后,MYC2转移到细胞核中,调节与JA反应相关的基因的表达 。 最终,MYC2作为一个关键调控因子,协调各种机制以减轻Cd胁迫引起的氧化损伤、提高抗氧化酶的产量、促进东南景天的生长并改善Cd的吸收和富集 (图10C)。
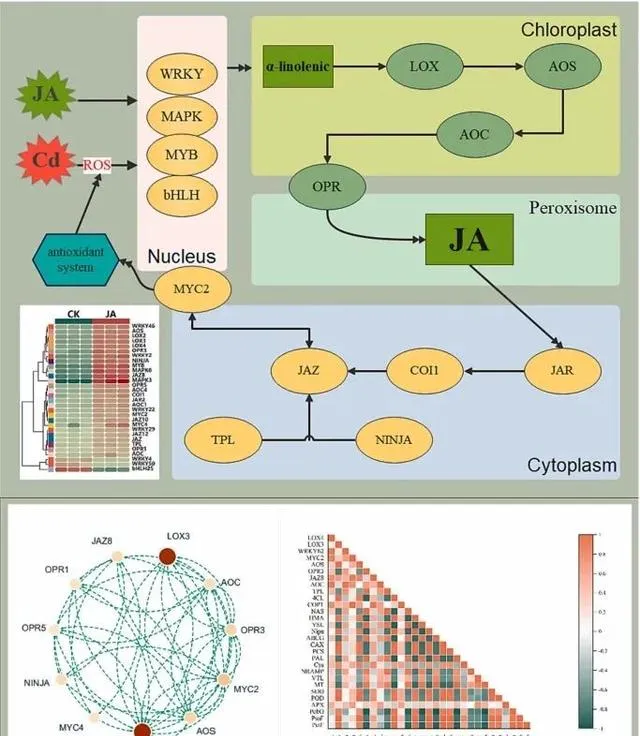
图10.JA在Cd胁迫中的调控作用。 (A)JA生物合成和信号传导机制;(B)JA通路的蛋白质相互作用网络;(C)热图显示了东南景天中与JA、光合作用、抗氧化剂和Cd吸收/积累有关的基因相关性。
结论
本研究通过比较分析外源JA处理对Cd胁迫下东南景天的生理效应和转录组效应,揭示了JA在调节植物对Cd胁迫的响应中发挥的多种作用 。 本研究结果表明,施用JA能够减轻Cd胁迫对植物生长的不利影响,通过优化光合作用促进正常的生理条件和生长 。此外, JA在增强植物抗氧化防御机制方面发挥了重要作用,通过调节编码抗氧化酶的基因的表达,帮助清除ROS,从而维护细胞内的稳定和秩序 。更重要的是, JA处理明显减少了Cd在植物体内的积累,表明JA有可能成为减轻Cd胁迫影响的有效策略 。 转录组分析发现了一系列在JA介导的Cd胁迫响应中起作用的DEGs, 为了解JA如何通过调控基因表达来操纵植物响应提供了新的视角。 它涵盖了编码参与抗氧化系统和光合作用功能的基因,并涉及细胞壁合成、金属螯合和转运等多个方面,展示了一个多层次、错综复杂的调控网络 。总之,本研究深入揭示了JA在东南景天的Cd胁迫响应中的作用,为其在植物生物技术和环境修复中的应用前景铺平了道路。











