文章1

低氧应答途径调控兰科植物铁皮石斛原球茎与真菌的共生
期刊:
IF: 7.4
发表时间: 2023年3月
单位: 云南大学
研究背景
云南大学生态与环境学院生物多样性研究院韩佳嘉和高江云团队在在线发表了题为「Symbiosis between protocorms and involves the plant hypoxia response pathway」的研究论文,该研究利用转录组,基因功能验证等方法揭示了真菌是一种能与多种兰科植物建立菌根共生关系的内生真菌,并发现植物ADH(乙醇脱氢酶)及其相关的低氧应答途径调控了兰科植物铁皮石斛原球茎与真菌的共生。 迈维代谢为其提供了转录组检测与分析服务。
菌根是真菌和植物根之间普遍存在的共生体。兰科植物种子细小,胚未完全分化、几乎没有胚乳,需要依靠特定的共生真菌提供营养来促进其萌发和后续原球茎的生长。不同于有广泛宿主范围的丛枝菌根真菌,兰科植物真菌对他们的宿主通常具有高度特异性。然而,与丛枝菌根和根瘤菌共生的分子机制相比,兰科菌根共生的分子机制尚不清楚。在本研究中,作者发现内生真菌是一种广谱的兰科植物亲和性真菌,能够促进多种(2个属6个种)附生兰科植物的种子萌发并促进其原球茎发育成幼苗,其中包括铁皮石斛()。铁皮石斛原球茎与真菌的共生,为后续研究提供了重要模型。
研究材料
12种生态和分类学上不同的兰科种子,无菌和共生铁皮石斛原球茎。
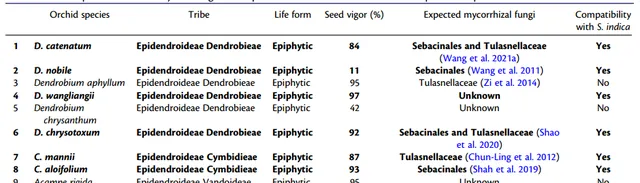
技术路线
研究结果
1、能够与铁皮石斛()及其他兰科植物共生
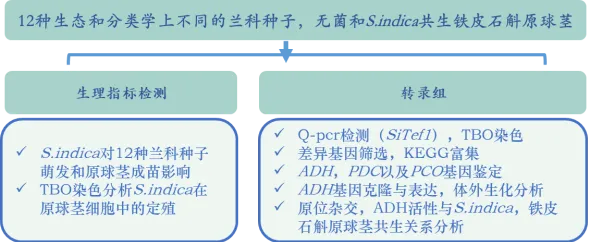
作者分析了内生真菌对收集的12种生态和分类学上不同的兰科种子萌发和原球茎成苗的影响。结果发现,促进了其中6种的生长:铁皮石斛()、金钗石斛()、王亮石斛()、鼓槌石斛()、纹瓣兰()和硬叶兰()(图1A,B)。不仅刺激了这些种子的萌发,而且促进了原球茎的发育(图1A,B)。对铁皮石斛的促生长作用较强,但对硬叶兰较弱(图1B)。作者通过半薄组织切片甲苯胺蓝(TBO)染色分析了在原球茎细胞中的定殖。细胞内菌丝体位于兰科植物原球茎的基部区域(图1C),该区域为未来根系发育的位置。细胞内菌丝团(图1C)在原球茎中的形态与其他兰科菌根(OrM)一致。因此,该研究结果表明,与一些兰科植物形成了典型的OrM,并促进了其种子萌发和原球茎的发育。在这些兰科植物中,铁皮石斛是一种具有较高药用价值的濒危植物,并且已经公布了参考基因组。因此作者利用铁皮石斛-共生系统研究了兰科植物与内生真菌之间的共生机制。
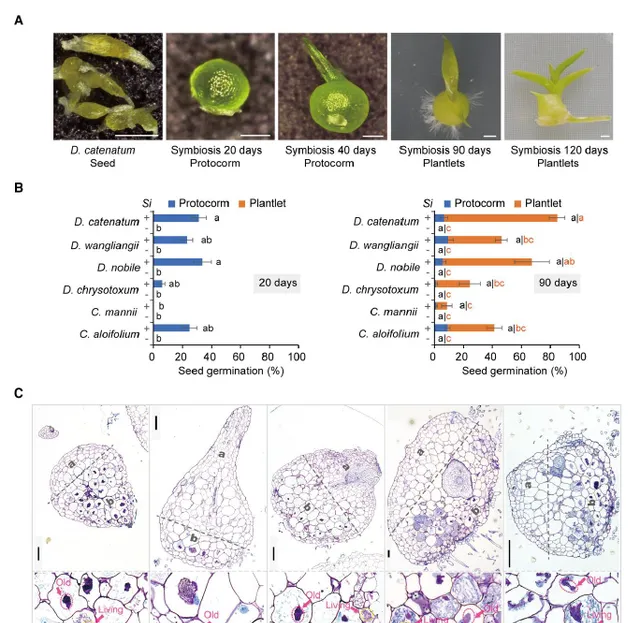
图1 与多种兰科植物共生。A:铁皮石斛种子和原球茎在接种后的形态;B:(Si)促进6中兰科植物种子萌发(接种约20d后形成原球茎)和原球茎发育(大多数原球茎在90天内发育成小植株);C:半薄组织切片甲苯胺蓝(TBO)染色的原球茎切片,(a)兰科原球茎的顶端未感染区和(b)基部菌根区,底部为定植细胞的细节。
2、共生原球茎与无菌原球茎的表达谱差异分析
在这项研究中,为了获得足够的样本量用于RT-qPCR和转录组分析,作者使用了无菌萌发种子30天龄的铁皮石斛原球茎作为原材料,将30天大的原球茎移植到共生培养基(OMA培养基)中进行无菌培养(CK)和接种(Si),然后收集用于随后的RT-qPCR和转录组分析。对在接种后的原球茎中的翻译延伸因子1()表达进行RT-qPCR分析,以铁皮石斛Actin-7()为内参,RT-qPCR结果显示定殖水平在接种后第9天达到峰值(图2A)。半薄组织切片甲苯胺蓝(TBO)染色结果也证实了共生原球茎中存在细胞内菌丝体,位于原球茎的基部区域(图2B),这与前期观察结果一致(图1C)。因此在接种后第9天的原球茎和真菌之间建立了稳定的共生关系。作者选择接种后第9天的共生原球茎用于随后的转录组分析。
转录组分析显示,无菌(control)和共生原球茎()的表达谱不同(图2C,D)。作者从vs control中筛选鉴定出356个差异表达基因(DEG)(log2(fold change)≥1,q≤0.05),KEGG通路富集发现,这些DEGs主要富集在植物防御和免疫应答相关通路(图2E),包括植物激素信号转导、植物病原体互作和MAPK信号通路,除此之外还包括类黄酮生物合成、苯丙素生物合成和色氨酸代谢通路。在大多数富集途径中,上调和下调的DEG数量相似(图2E)。观察到第18天的定殖水平低于第9天(图2A),作者认为在稳定的菌根共生关系建立后,菌根真菌在寄主植物中的定殖是有限的,并且这种定殖影响了兰科植物的防御和免疫相关途径。此外,糖酵解途径4个基因在糖酵解途径中上调,推测ADH在铁皮石斛菌根共生中起着重要作用。
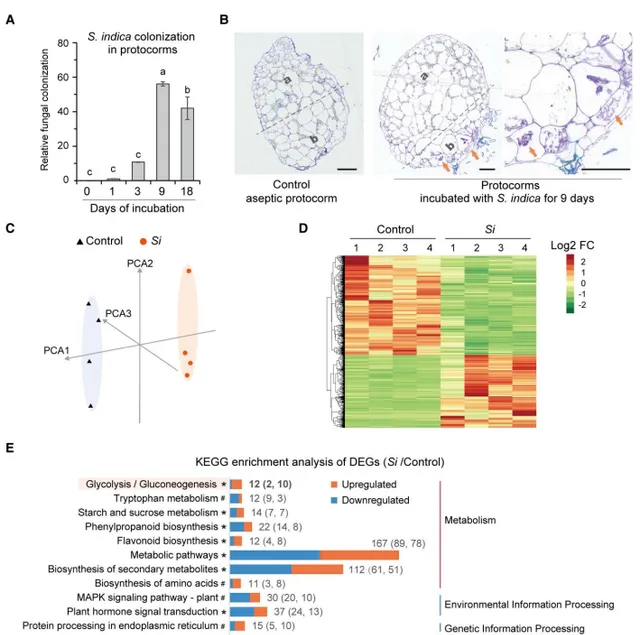
图2 共生原球茎转录组分析。(A)的RT-qPCR分析;(B)无菌和共生原球茎顶端区域(a)和基底区域(b)的半薄组织切片;(C)转录组数据PCA分析;(D)聚类热图分析;(E)差异表达基因(DEGs)的KEGG富集分析。
3、低氧适应性基因在共生原球茎中被诱导
ADH和PDC(丙酮酸脱氢酶)在植物低氧反应过程中发挥着重要作用。通过同源性分析,作者鉴定出12个同源基因。转录组和RT-qPCR分析显示,在共生原球茎中,几乎所有的基因都被上调,其中和上调最显著(图3A,C)。生化分析表明接种后第9天,共生原球茎中ADH的还原活性明显高于无菌原球茎,与上述结果一致(图3D,E)。这些结果表明,和可能与菌根共生特异性相关。此外,基因在共生原球茎中显著上调(图3B,C)。缺氧诱导基因和也上调(图3B,C)。此外,和等低氧适应性基因的表达与共生时间呈正相关。这些结果表明,的定植能够诱导铁皮石斛共生原球茎中低氧适应性基因(特别)的表达。
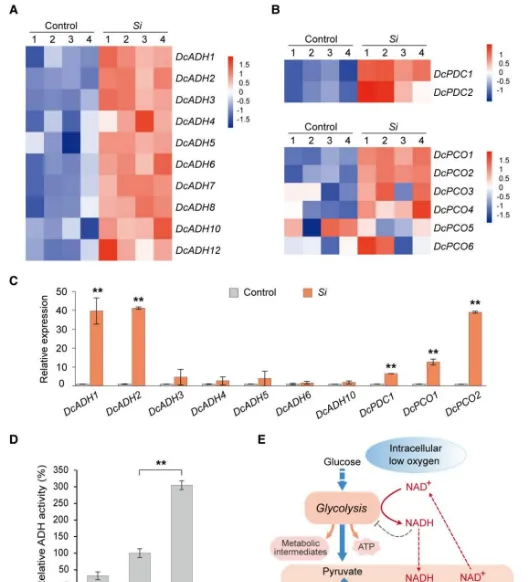
图3低氧适应性基因在共生原球茎中被诱导。(A、B)和基因表达热图;(C)关键的低氧适应性基因的RT-qPCR分析;(D)无菌原球茎以及共生原球茎中ADH活性;(E)植物缺氧代谢受ADH和PDC调控。
4、ADH活性正向调节原球茎菌根共生
为了验证的上调是否与共生原球茎中ADH活性升高有关,作者克隆了和的编码序列,并在大肠杆菌中表达。重组菌蛋白提取物的体外生化分析显示,和具有预期的ADH活性,可将NADH转化为NAD+(图3E和4A)。接着作者证实了葛根素和4-甲基吡唑(ADH抑制剂 )均抑制了和的NADH转化活性(图4B,C)。其中在共生原球茎中表达量最高。因此作者使用地高辛(DIG)标记的寡核苷酸探针进行了原位杂交。在共生原球茎中,主要在菌根基部表达,而不是在根尖表达(图5A,B),这与在原球茎中的定殖位置一致。接着作者使用葛根素来抑制的功能,并观察其对与铁皮石斛原球茎共生关系的影响。作者发现葛根素显著降低了在铁皮石斛原球茎上的定植(图5C),但对的生长没有显著的不利影响(图5D)。另一种ADH抑制剂4-甲基吡唑也显著降低了在铁皮石斛原球茎上的定植,但在200 μM浓度下,4-甲基吡唑能轻微抑制的生长。这些结果表明,抑制ADH活性可以抑制在铁皮石斛原球茎上的定植。综上结果表明,ADH及其相关的低氧应答途径调控兰科植物铁皮石斛原球茎与真菌的共生。
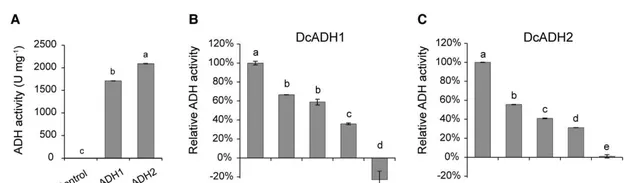
图4 和蛋白的ADH活性。(A)体外实验表明,纯化后的和蛋白具有ADH还原活性,可催化NADH转化为NAD+;(B, C)不同浓度葛根素对纯化后的和还原活性的抑制作用。

图5 ADH活性是共生原球茎的OrM真菌定殖所必需的。(A) mRNA在无菌和共生原球茎的纵向切片上的原位杂交;(B)正义(而非反义)探针的原位杂交;(C)体内试验表明,在接种后第9天,葛根素抑制了在共生原球茎中的定殖;(D)葛根素对的生长没有产生显著的不利影响;(E)ADH调节兰科原球茎中菌根共生的模式图。
文章2

转录组+代谢组解析人参皂苷胁迫诱导三七根分泌物变化以招募有益的伯克霍氏菌株B36
期刊: J. Agric. Food Chem.
IF: 6.1
发表时间: 2023年3月
单位: 云南农业大学
研究背景
云南农业大学植保学院杨敏老师团队在J. Agric. Food Chem.在线发表了题为「Autotoxic Ginsenoside Stress Induces Changes in Root Exudates to Recruit the Beneficial Strain B36 as Revealed by Transcriptomic and Metabolomic Approaches」的研究论文,该研究利用转录组和代谢组等方法阐明了人参皂苷胁迫激活了苯丙素生物合成和α-亚麻酸代谢通路,从而增加了肉桂酸、2-十二烯酸和12-氧代植物二烯酸(12-OPDA)的分泌。其中肉桂酸可以同时促进B36的趋化和生长,增强B36在根际定殖,最终提高三七的成活率。这一发现将促进有益菌在农业生产中的实际应用,并通过外源添加关键代谢产物实现可复制的生物防治效果。 迈维代谢为其提供了代谢组和转录组检测分析服务。
三七((Burk.)F.H.Chen)由于土传病原菌和自毒性物质人参皂苷的积累,易患严重的根腐病。前期研究表明,在人参皂苷自毒胁迫下,三七能在根际土壤中富集有益的伯克霍尔德菌B36,该菌能降解自毒皂苷 同时拮抗土传病原菌 ,最终缓解根腐病,这为通过控制根际微生物来克服根腐病提供了一种新的生态策略。然而,自毒性物质人参皂苷是否能诱导三七根系转录组和分泌物的变化,进而募集有益微生物,目前尚不清楚。
研究材料
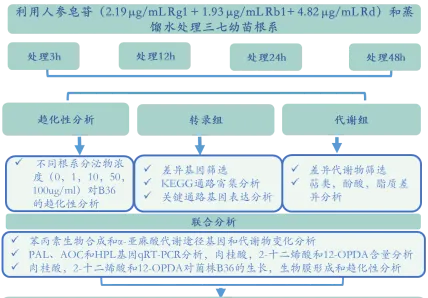
三七幼苗在人参皂苷(2.19 μg/mL Rg + 1.93 μg/mL Rb+ 4.82 μg/mL Rd)溶液和蒸馏水溶液中培养3、12、24和48h,在每个时间点下收集根组织进行转录组分析并收集玻璃瓶中的溶液,用滤纸和0.22 μm亲水性膜过滤后浓缩成干物质进行代谢组分析。每个处理3个生物学重复。
技术路线
研究结果
1、人参皂苷胁迫下三七根系分泌物可吸引伯克霍尔德菌B36
当三七根受到人参皂苷胁迫后,三七根系分泌物对B36表现出明显的吸引作用,且呈剂量依赖性(图1))。具体而言,当分别利用胁迫处理3h,24 h下浓度为10、50和100 μg/mL的三七根系分泌物浓度以及12 h下浓度为100 μg/mL处理时对菌株B36表现出显著的吸引作用(图1A,B,C)。人参皂苷胁迫48 h后,与无菌水对照相比,根分泌浓度为50 μg/ mL时对菌株B36具有显著的吸引作用(图1D)。此外,人参皂苷灭菌水处理与对照之间无显著差异。上述结果表明,在人参皂苷胁迫下,三七根系可以分泌一些代谢产物来吸引B36。
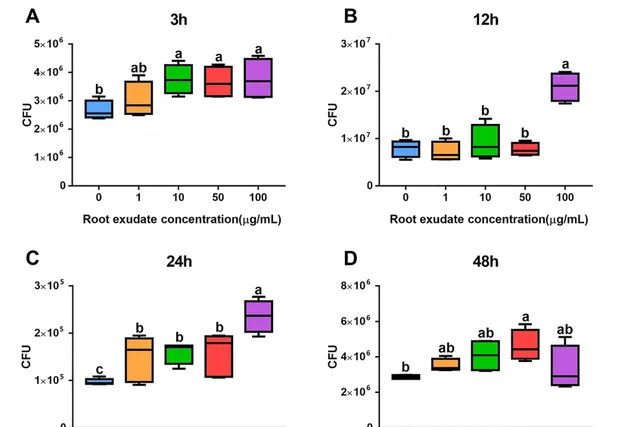
图1 人参皂苷胁迫处理3 (A)、12 (B)、24 (C)、48 h (D)时不同浓度根分泌物对伯克霍尔德菌B36的趋化性测定。
2、人参皂苷胁迫处理改变三七根的转录组谱
利用人参皂苷和蒸馏水分别胁迫处理3、12、24和48 h的三七根进行转录组分析。共24个文库(ck-3h, g-3h, ck-12h, g-12h, ck-24h, g-24h, ck-48h, g-48h,每个处理3个生物重复)得到约159.62 Gb的clean bases,Q20和Q30的平均值分别为98.11和94.32%,平均GC含量为43.66%。利用NR、GO、KEGG、Pfam、KOG和SwissProt数据库注释后进行差异基因筛选。4个比较(ck-3h .g-3h, ck-12h .g-12h, ck-24h .g-24h, ck-48h .g-48h)分别筛选到33、15、713和126个DEG。其中分别有25、5、615和106个DEG在人参皂苷处理时显著上调(图2A)。说明人参皂苷胁迫24 h后,根内基因表达水平发生了显著变化。KEGG通路富集分析结果显示,在人参皂苷胁迫3、12、24和48 h后,分别有1、1、9和2个通路被显著富集(图2B),其中苯丙素生物合成途径在胁迫3和12 h下均显著富集(图1)。在胁迫24 h后显著富集的9条途径中(图2B),苯丙素生物合成和二芳基庚烷和姜醇生物合成途径的DEGs大部分上调表达(图2C)。在苯丙素生物合成过程中,大部分编码过氧化物酶(POD)、反式肉桂酸4-单加氧酶(CYP73A)、β-葡萄糖苷酶、反式肉桂酸-4-羟化酶(CYP98A)、莽草酸羟基肉桂酰基转移酶(HCT)、肉桂醇脱氢酶(CAD)和东莨菪素葡萄糖转移酶(TOGT1)的基因在人参皂苷处理24 h后显著上调,其中编码CYP73A和CYP98A的基因(Pno05G006544和Pno01G007403)在人参皂苷处理24 h后分别上调了2.99倍和3.43倍。二芳基庚烷和姜醇生物合成中编码反式白藜芦醇二-O-甲基转移酶(ROMT)的基因也显著上调。另外在富集到α-亚麻酸代谢、花生四烯酸代谢及其上游途径不饱和脂肪酸生物合成和脂肪酸代谢中的DEGs均上调(图2C)。例如编码FAD2、ACSL、TGL4、CYP4A、DAD1、OPR、DOX、HPL的基因。其中,编码OPR的基因(Pno02G004829和Pno02G004840)在人参皂苷处理24 h后分别上调了3.48倍和3.47倍。这些结果表明,参与上述途径的DEG可能与三七根对人参皂苷胁迫的反应密切相关。
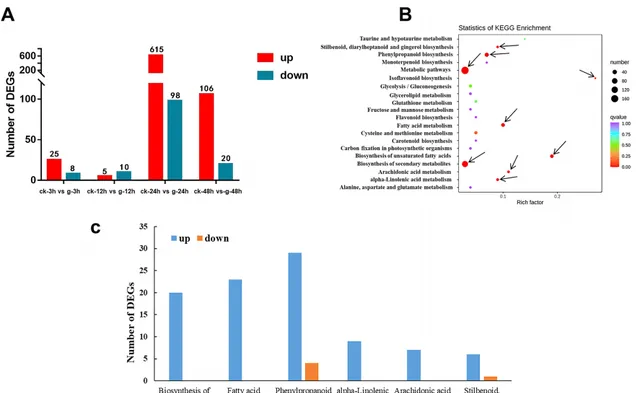
图2 人参皂苷胁迫对三七根转录组影响。(A)上调和下调基因柱状图统计;(B)人参皂苷处理24 h后KEGG富集分析;(C)人参皂苷处理24 h后显著富集途径中的DEGs数量。
3、人参皂苷胁迫下根系分泌物代谢组学分析
在所有样品中总共检测到273种代谢物,PCA结果显示胁迫不同时期根系分泌物代谢谱与对照组差异显著,且不同时期间也明显分离。通过VIP≥1和|log2(FC)|≥1筛选差异代谢物(DAM),共得到250个DAM,其中人参皂苷胁迫处理3、12、24、48 h时,代谢物分别上调34、31、28、22个,下调52、27、41、15个。根分泌物中响应人参皂苷胁迫的DAM主要分为7类,包括氨基酸及其衍生物、酚酸、核苷酸及其衍生物、萜类、有机酸、脂类等(图3A)。在人参皂苷处理3、12和24h后,萜类化合物(人参皂苷Rg1、人参皂苷Rp、三七皂苷A4、三七皂苷R1和三七皂苷R2)均显著上调(图3A)。此外,人参皂苷处理24h后,酚酸(肉桂酸、4-甲氧基肉桂醛、2氨基-3-甲氧基苯甲酸等)和脂质(2-十二烯酸、12-OPDA等)显著上调,而有机酸(2-氧庚二酸、2-甲基-3-氧己二酸等)显著下调(图3B)。因此作者推测人参皂苷胁迫可能导致三七根分泌物中萜类、酚酸和脂类积累增加。
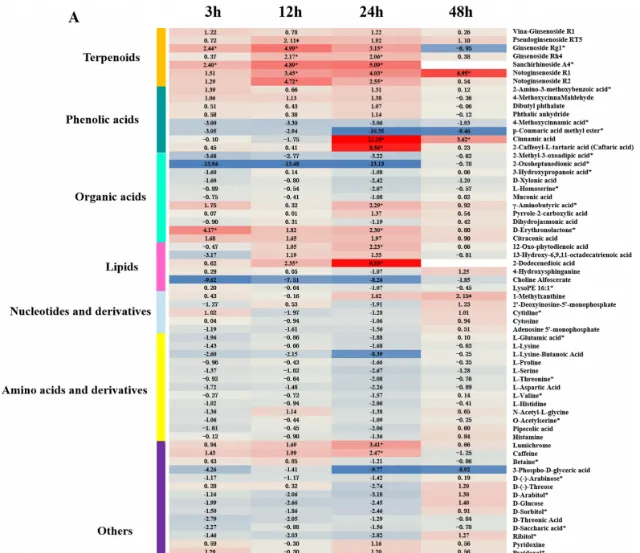
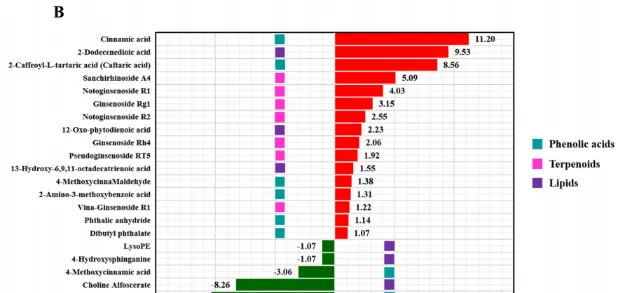
图3 人参皂苷胁迫下代谢物差异分析。(A)人参皂苷处理不同时期代谢物显著变化热图;(B)人参皂苷处理24 h后酚酸、萜类和脂类变化条形图。
通过结合转录组和代谢组联合分析(图4A),作者选择苯丙素生物合成和α-亚麻酸代谢途径中显著变化的基因,包括编码PAL、AOC和HPL的基因进行qRT-PCR分析(图4B)。同时作者检测了人参皂苷处理24h后肉桂酸,2-十二烯酸和12-OPDA含量变化,发现处理后三七根分泌物中肉桂酸含量显著增加至2511.194 ng/g,2-十二烯酸和12-OPDA含量显著降低(图4C)。
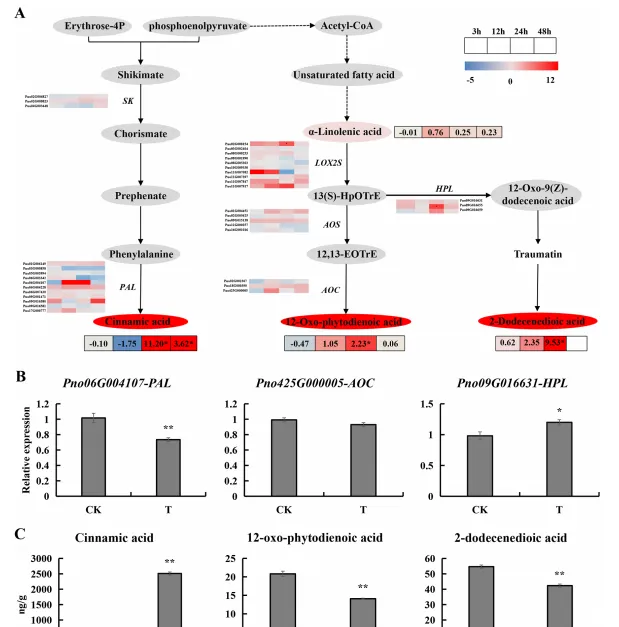
图4 人参皂苷胁迫3、12、24和48 h后苯丙素生物合成和α-亚麻酸代谢相关基因和代谢物表达谱;(A)苯丙素生物合成和α-亚麻酸代谢通路热图;(B)PAL、AOC和HPL基因表达量;(C)肉桂酸、2-十二烯二酸和12-OPDA含量。
4 人参皂苷刺激三七根系分泌物中代谢物积累以增强B36定植
研究发现,人参皂苷胁迫促进三七根中肉桂酸分泌增加。趋化性实验数据显示,低浓度(1、10和50 μg/mL)的肉桂酸对菌株B36有明显的吸引作用(图5A),且在10和100 μg/mL浓度下显著促进了菌株B36生物膜的形成(图6A),在50μg/mL浓度下培养36 h时显著促进了菌株B36细胞的生长(图6D)。然而,高浓度的2-十二烯酸和12-OPDA(100 μg/mL)对菌株B36具有驱避性(图5B,C)。2-十二烯酸对生物膜的形成没有明显的促进作用(图6B)。这些结果表明,肉桂酸可能是根际B36趋化的重要信号物质。
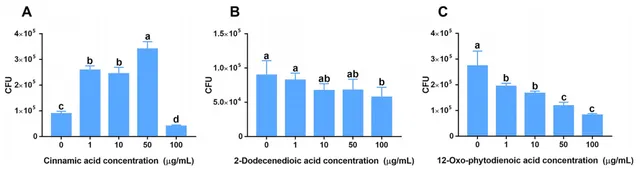
图5伯克霍尔德菌B36对肉桂酸(A)、2-十二烯二酸(B)和12-OPDA (C)的趋化反应。
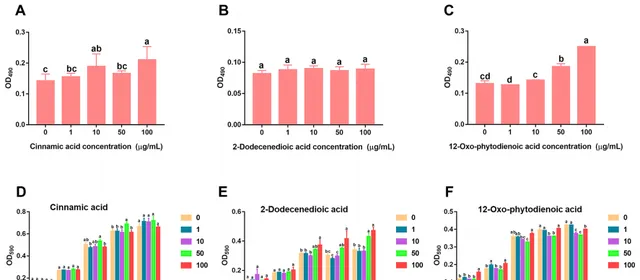
图6 肉桂酸、2-十二烯酸和12-氧-植物二烯2-十二烯二酸(B)和12-OPDA对伯克霍尔德菌B36生物膜形成(A−C)和生长(D−F)的影响。
结合以上结果,作者进一步检测了肉桂酸对B36在无菌土壤中定殖能力的影响。利用三七根分泌物和肉桂酸处理B36土壤,在处理3、5、10 d后,B36的定殖能力显著增强。更重要的是,与根系分泌物处理相比,肉桂酸处理下B36在无菌土壤中的定殖数量显著增加(图7A)。盆栽试验进一步证实,在连作土壤中单独添加肉桂酸和B36后,对三七的存活率并没有明显的促进作用,但肉桂酸与B36配合使用,可以显著提高三七在连作土壤中的存活率(图7B)。
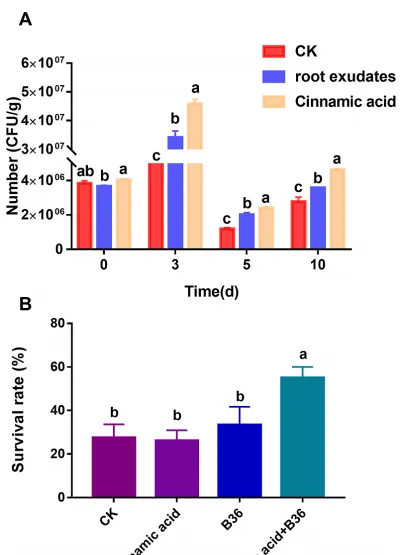
图7 三七根分泌物和肉桂酸对无菌土壤中B36定植的影响(A),肉桂酸和B36对连作土壤中三七成活率的协同效应(B)。
往期热门分享














