
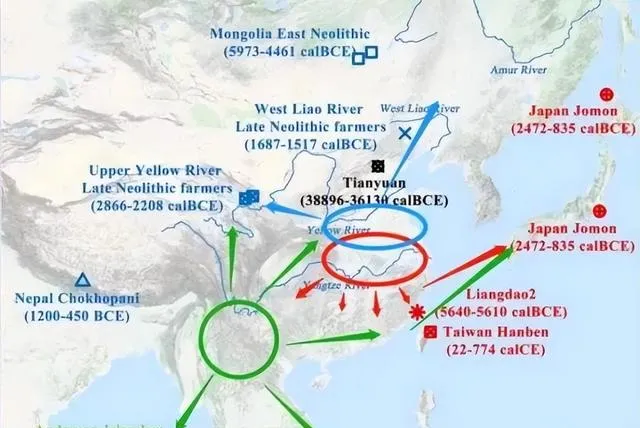
關於東亞父系NO起源問題,多數人認同西南起源說,但相當部份人認為西北/北方起源說。雙方各持一辭,似各有道理,但事實真的如此嗎?隨著隨著古DNA提取及測序技術不斷完善,南北各地古DNA數據量越來越大,進一步深化了我們對祖先起源的認識。現有的古DNA數據,與西南起源說、西域起源說有很大的出入。根據考古調查報告,新石器時代早期中期華北地區及其山東一帶發現了很多N-M231,晚期開始出現了O-M175,南方新石器中期至末期出現了很多O-M175、但N-M231極為稀缺。已知K2a2b(NO)-M214有兩大分支:N-M231與O-M175。如果哪些地區N-M231與O-M175交替共存,下遊分支齊全,那麽最可能是NO-M214發源地,向周邊地區擴張。
在黃河中下遊,N-M231的下遊分支豐富多樣,包括N1-CTS3750與N2-B482。其中N1-CTS3750衍生了兩大分支N1a-F1206與N1b-F2930。N2-B482集中分布於中國華北、東歐。
N1a-F1206:東歐、西伯利亞、中國北方,但N1b-F2930稀缺。
N1b-F2930:中國東部、藏緬語族、中南半島,但N1a-F1206稀缺。
N-M231的下遊分支(N1a、N1b、N2)區域交集,結果為黃河中下遊,可以推測黃河中下遊最可能是N-M231發源地,粟作農業文化人群與N-M231具有密切相關性。
O-M175有兩大分支:O1-F265與O2-M122。其中,O1-F265衍生了兩大分支:O1a-M119與O1b-M268。
O1a-M119:中國南方、東南亞。
O1b-M268衍生了兩大分支:東南亞O1b1-F2320與日韓O1b2-P49
O2-M122衍生了兩大分支:東亞/太平洋O2a-M324與華東O2b-F742
O-M175的下遊分支(O1a-M119、O1b-M268、O2-M122)區域交集,結果為東亞區域,可以推測O-M175最早分化地點最可能是中國東部,稻作農業文化人群與O-M175具有密切相關性。
不難看出:在黃淮海平原(華北與淮河之間),N-M231與O-M175交替共存,下遊分支齊全,那麽最可能是NO-M214發源地。

距今六千~五千年的魯北大汶口文化傅家遺址男性則有多樣化的Y染色體序列,實際測到K*(×O)有一例、N為兩例、O為一例。同期河北紅山文化姜家梁遺址全是12例N。由於魯北與河北為鄰,,必有人員流動,出現了N、O並存的現象,清晰表明大汶口文化北上擴張的過程,進一步證明新石器時代山東地區N-M231與O-M175交替共存,最可能是NO-M214發源地。
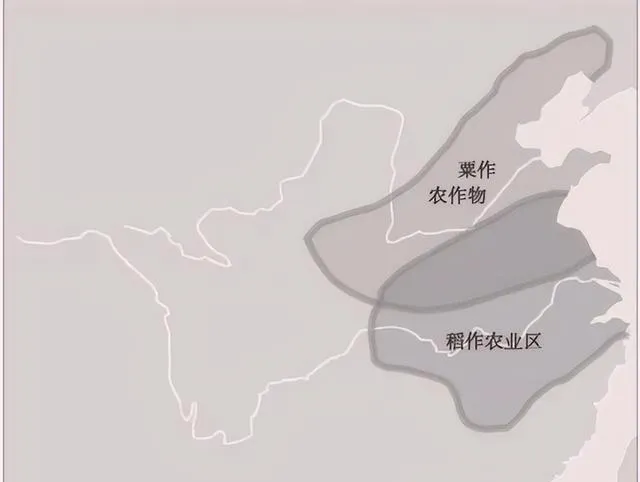
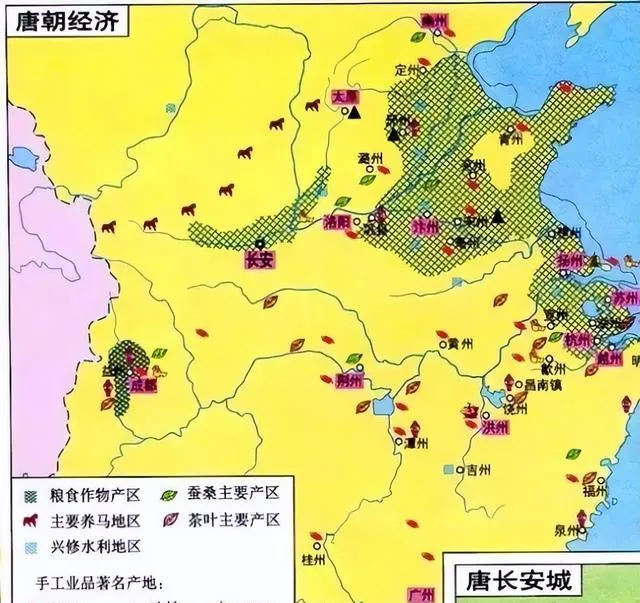
此圖出自農業部。古代主要糧食作物產區地幾乎都在黃河中下流域,而西北地區、北方地區都是遊牧民族的主要養馬地區,對於東方農業文明構成嚴重的威脅。
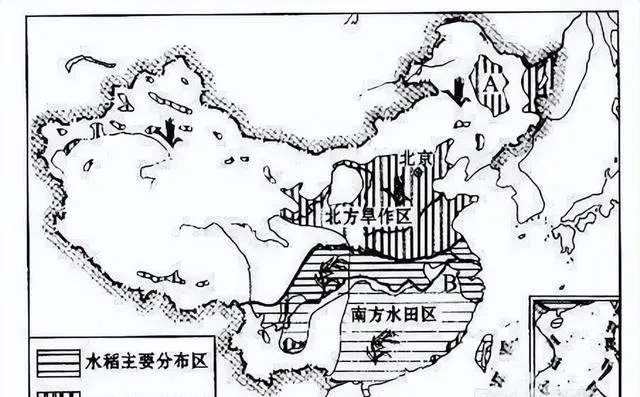
N系人群率先到達黃河流域北岸,開創了粟作農業,但生產規模不大,至今還沒有成為中國人的主糧,N系人群漸漸地結束歷史舞台,但留下一定的文化遺產。
O2-M122早在3萬年前就已到達東亞,掌握了農業種植技術,在來東亞之後才開始其新石器時代的擴張,與中國農業的東部本土起源是一致的。總之,O2-M122強烈人口擴張的Y染色體支系在中國東部的廣泛分布,由於中國東部大平原屬半濕潤、半幹旱氣候地區,當初從事稻粟混作,後來引進了小麥,致力於在中原地區推廣小麥種植,奠定了黃河農業文明的基礎,小麥最終稱為北方漢人的主糧。東南亞O1a-M119和O1b1a1-M95長期生活在熱帶森林從事半農半獵,遠不如黃河農業文明高度發達。湖南、浙江一帶最早出現水稻,但生產規模不大,也不是商周秦漢的主糧。自從漢代統一南方,才開始開拓南方農業,稻栽培已具相當規模。自從宋代以來,水稻慢慢變成了多數中國人的主糧。

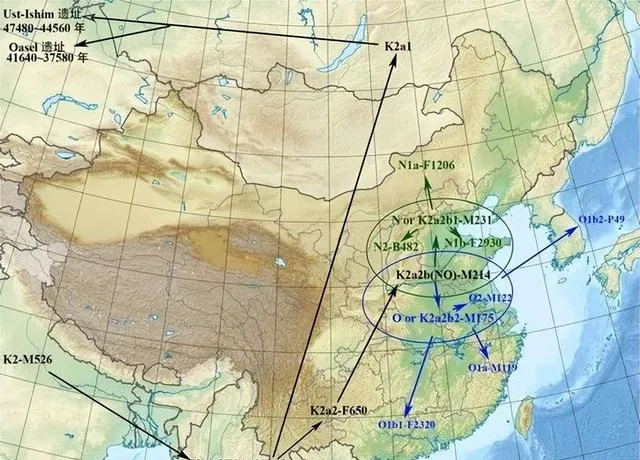
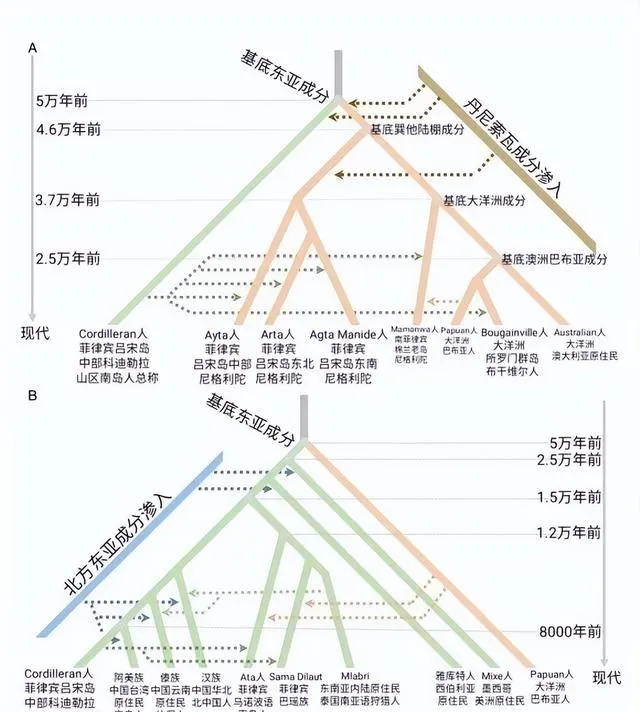
Ust-Ishim人(烏斯特-伊斯姆人是K2A)是在俄羅斯西伯利亞西北地區發現的4.5萬年前的Y染色體為k2a的人類遺骨。K2a 或者說pre NO是NO的祖形(未分化),和其他的K2支系相比分化較早, 其他的K2支系包括K2b、K2c、K2d、K2e。K2a 最早與其他K2集團分離,在印度巴基斯坦地帶向北遷徙, 其他K2則繼續挺進南亞、東南亞、澳洲。所以,東南亞K2多樣性很高,推測K2a-CTS11667最可能起源於南線,估五萬年前到達東亞沿海,沒有繼承北歐亞ANE人群的一丁點常染,但是繼承了田園洞人的東歐亞常染。田園洞人或者Ust-Ishim人是東歐亞、南歐亞、巴布亞澳洲共同的一個祖先,並且現代東亞NO人群不含ANE常染成分,否定了NO西北/北方起源說。


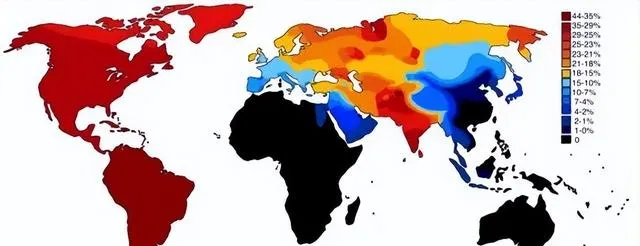
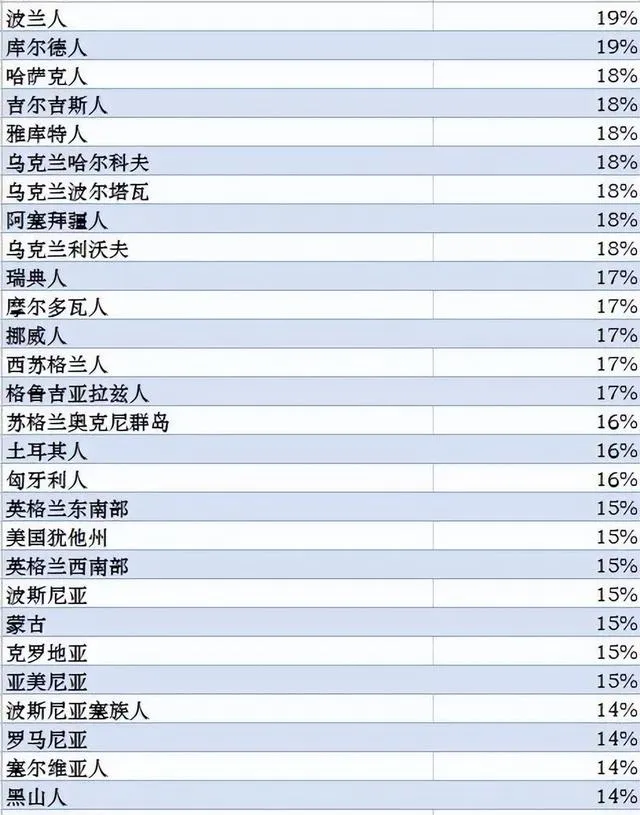
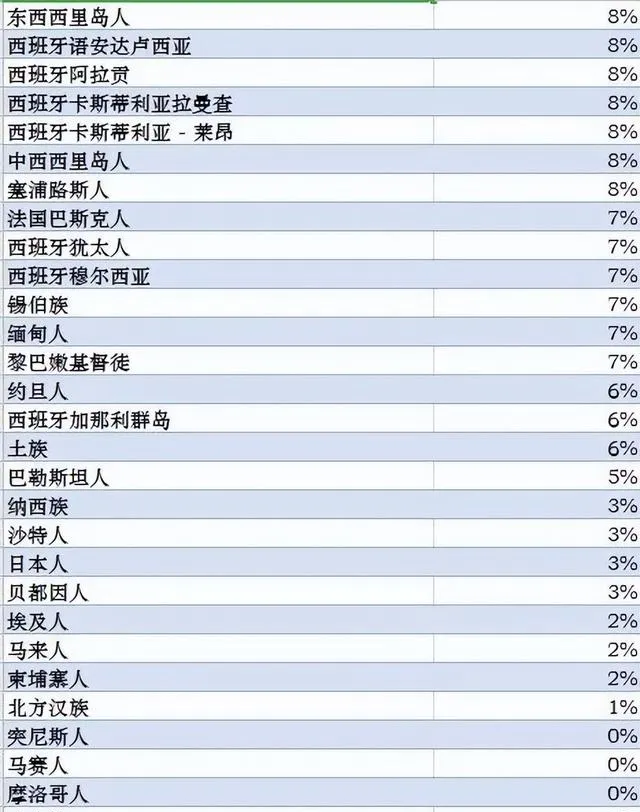
世界各族ANE血統比例分布圖,ANE古北歐亞人是大概兩萬年前中石器時代分布在歐亞大陸北部的一個人群,可以解析為70%的西歐亞和30%的東歐亞血統的混合。參考此數據制作而成。




首先要了解一下現代歐洲人形成的過程。7萬年前智人走出非洲,在中東混入了少量尼安德特人基因,然後分離成東歐亞人(East Eurasian)、西歐亞人(West Eurasian)這兩大分支。有一支沒混入尼安德特人基因的人群叫Basal Eurasian留在了中東和北非。由於沒有混入尼人基因,Basal Eurasian比較接近撒哈拉以南的非洲人。
東歐亞人東遷後分離成形成東亞人種、大洋洲土著人種。
而西歐亞人遷徙到歐洲,形成西歐亞狩獵采集者(WHG)。另一種西歐亞人遷徙到西伯利亞,混入了30%~20%的東歐亞基因形成古北歐亞人(ANE),之所以現代西伯利亞土著不是高加索人種是因為後來東亞人種不斷北上遷徙稀釋當地ANE血統的結果。ANE對後期草原牧民古印歐人有很大的血統貢獻,後來印歐人大規模遷徙到歐洲。新石器時代一支西歐亞人和中東的Basal Eurasian人混合形成中東新石器時代農民(EEF),EEF也大規模遷徙到歐洲,並帶來農業種植技術。
現代歐洲人正是西歐亞狩獵采集者(WHG)、古北歐亞人(ANE)、Basal Eurasian這三種遺傳差異較大的人群混合的結果。根據對歐洲古人類基因的檢測,現西歐亞狩獵采集者WHG人群大多同時具有深色皮膚、藍眼睛、黑頭發的特征,但遺傳特征接近現代北歐人。


付巧妹團隊研究觀察到,現今的東亞人群,不管是來自北方還是南方,都同時混合有古北方人群成分(以黃河沿岸的山東人群為代表)和古南方人群成分(以福建與台灣海峽島嶼人群為代表),但古北方人群成分影響更為廣泛(圖2C和圖3C)。而與福建奇和洞人相關的古南方人群成分在中國南方大陸大量下降(圖3C和3F)。該結果表明人群的遷移,包括新石器時代南北方人群之間的互動融合,以及 新石器時代以後黃河以北人群向南的大量遷移,基本構成了現今東亞南北方人群的基因組成。
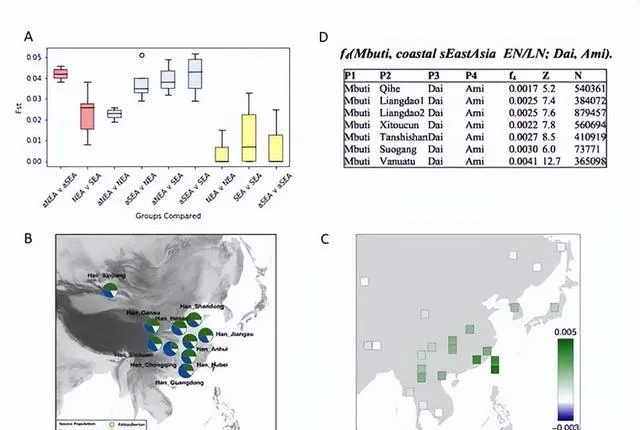
(A)不同人群的遺傳分化指數分布圖,指數越高表示相關人群的分化程度越高,遺傳差異越大,該圖表明新石器時代南北方人群的的遺傳差異遠高於現今東亞南北方人群。(B)現代漢族人群祖源成分與地理分布圖。 現今中國南北方的漢族人群,不管是在南方還是北方,均顯示混合有東亞古南方人群成分、古北方人群成分、古西伯利亞人成分。其中,現代漢族人群裏攜有古北方人群成分比例最高的群體顯示位於山東,這裏也正是本研究古北方人群的代表個體樣本的遺址所在地,由此反映出山東地區人群自新石器時代以來的遺傳連續性。 (C)現今東亞人群與古南方人群、北方達斡爾族人群的遺傳關系圖,綠色表示與古南方人群的關系更近,藍色表示與達斡爾族人群更近。(D)古南方人群與現今傣族人群、台灣島阿美族人群的遺傳關系的f4分析圖,Z>3 表示與阿美族人群關系更近。

復旦倫敦新論文中的截圖譯文:(圖4B)進行的成對遺傳同質性的檢驗結果證實,漢代個體(受降城漢軍烈士、大槽子漢墓)與新石器早期山東個體的遺傳同質性高於黃河內陸(仰韶組和齊家組),漢代個體在PCA(圖1A)和qpAdm(圖3B)中的位置更接近於新石器時代的黃河先民。這表明,黃河內陸與沿海的混合在北方漢人的形成中起著重要作用。
這個樹圖出自2021年3月發表的【全基因組見證河西走廊人群混合史】論文,明確指出:九千年前山東小高遺址是100%東亞人後裔。北方粟作農人是由68%東亞成分和 32%類Onge成分混合的,仰韶文化汪溝遺址是北方粟作農人的直屬後裔。南方稻作農人是由77%類Onge成分何23%東亞成分混合的,台灣漢本遺址是南方稻作農人的直屬後裔。甘肅蘭州漢族是由97%北方粟作農人和3%南方稻作農人混合的。
研究發現,廣西地區距今約1500年至500年的古人群與距今6000年的史前古老人群(包括廣西隆林人、獨山人和寶劍山人)完全不同,這意味著廣西特有的以隆林人為代表的相關古老東亞人群沒有任何基因貢獻,遠古廣西人徹底滅絕了。古代廣西人群更替,見證古東亞北方人群的擴張之道。九千年前古山東常染(古東亞北方人群),最終成為北漢血統主體。這一事實清楚告訴:K2a祖先五萬年前到達東亞沿海後所形成的東歐亞常染成分,作為東亞人種的奠基者,與後期南方的南歐亞成分(澳棕類別、和平文化人群)完全不同。
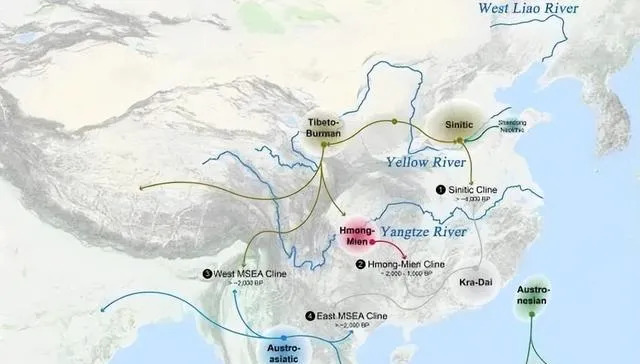
下面的祖先遷徙路線圖,又簡單又易懂。
原始東亞人(東亞農業人群)起源於黃淮海大平原,向西北/西南、華北/北亞、華南/東南亞、東北亞/日韓遷徙擴散,與當地土著遊牧人群或漁獵人群混血,形成了亞洲各族。
首先要了解一下現代歐洲人形成的過程。7萬年前智人走出非洲,在中東混入了少量尼安德特人基因,然後分離成東歐亞人(East Eurasian)、西歐亞人(West Eurasian)這兩大分支。有一支沒混入尼安德特人基因的人群叫Basal Eurasian留在了中東和北非。由於沒有混入尼人基因,Basal Eurasian比較接近撒哈拉以南的非洲人。
東歐亞人東遷後分離成形成東亞人種、大洋洲土著人種。
而西歐亞人遷徙到歐洲,形成西歐亞狩獵采集者(WHG)。另一種西歐亞人遷徙到西伯利亞,混入了30%~20%的東歐亞基因形成古北歐亞人(ANE),之所以現代西伯利亞土著不是高加索人種是因為後來東亞人種不斷北上遷徙稀釋當地ANE血統的結果。ANE對後期草原牧民古印歐人有很大的血統貢獻,後來印歐人大規模遷徙到歐洲。新石器時代一支西歐亞人和中東的Basal Eurasian人混合形成中東新石器時代農民(EEF),EEF也大規模遷徙到歐洲,並帶來農業種植技術。
現代歐洲人正是西歐亞狩獵采集者(WHG)、古北歐亞人(ANE)、Basal Eurasian這三種遺傳差異較大的人群混合的結果。根據對歐洲古人類基因的檢測,現西歐亞狩獵采集者WHG人群大多同時具有深色皮膚、藍眼睛、黑頭發的特征,但遺傳特征接近現代北歐人。
付巧妹團隊研究觀察到,現今的東亞人群,不管是來自北方還是南方,都同時混合有古北方人群成分(以黃河沿岸的山東人群為代表)和古南方人群成分(以福建與台灣海峽島嶼人群為代表),但古北方人群成分影響更為廣泛(圖2C和圖3C)。而與福建奇和洞人相關的古南方人群成分在中國南方大陸大量下降(圖3C和3F)。該結果表明人群的遷移,包括新石器時代南北方人群之間的互動融合,以及新石器時代以後黃河以北人群向南的大量遷移,基本構成了現今東亞南北方人群的基因組成。
(A)不同人群的遺傳分化指數分布圖,指數越高表示相關人群的分化程度越高,遺傳差異越大,該圖表明新石器時代南北方人群的的遺傳差異遠高於現今東亞南北方人群。(B)現代漢族人群祖源成分與地理分布圖。現今中國南北方的漢族人群,不管是在南方還是北方,均顯示混合有東亞古南方人群成分、古北方人群成分、古西伯利亞人成分。其中,現代漢族人群裏攜有古北方人群成分比例最高的群體顯示位於山東,這裏也正是本研究古北方人群的代表個體樣本的遺址所在地,由此反映出山東地區人群自新石器時代以來的遺傳連續性。(C)現今東亞人群與古南方人群、北方達斡爾族人群的遺傳關系圖,綠色表示與古南方人群的關系更近,藍色表示與達斡爾族人群更近。(D)古南方人群與現今傣族人群、台灣島阿美族人群的遺傳關系的f4分析圖,Z>3 表示與阿美族人群關系更近。
復旦倫敦新論文中的截圖譯文:(圖4B)進行的成對遺傳同質性的檢驗結果證實,漢代個體(受降城漢軍烈士、大槽子漢墓)與新石器早期山東個體的遺傳同質性高於黃河內陸(仰韶組和齊家組),漢代個體在PCA(圖1A)和qpAdm(圖3B)中的位置更接近於新石器時代的黃河先民。這表明,黃河內陸與沿海的混合在北方漢人的形成中起著重要作用。
這個樹圖出自2021年3月發表的【全基因組見證河西走廊人群混合史】論文,明確指出:九千年前山東小高遺址是100%東亞人後裔。北方粟作農人是由68%東亞成分和 32%類Onge成分混合的,仰韶文化汪溝遺址是北方粟作農人的直屬後裔。南方稻作農人是由77%類Onge成分何23%東亞成分混合的,台灣漢本遺址是南方稻作農人的直屬後裔。甘肅蘭州漢族是由97%北方粟作農人和3%南方稻作農人混合的。
付巧妹團隊研究發現,廣西地區距今約1500年至500年的古人群與距今6000年的史前古老人群(包括廣西隆林人、獨山人和寶劍山人)完全不同,這意味著廣西特有的以隆林人為代表的相關古老東亞人群沒有任何基因貢獻,遠古廣西人徹底滅絕了。古代廣西人群更替,見證古東亞北方人群的擴張之道。九千年前古山東常染(古東亞北方人群),最終成為北漢血統主體。這一事實清楚告訴:K2a祖先五萬年前到達東亞沿海後所形成的東歐亞常染成分,作為東亞人種的奠基者,與後期南方的南歐亞成分(澳棕類別、和平文化人群)完全不同。











