

鹿角蕨多組學的晝夜動態為弱CAM光合作用提供了新的見解
發表單位: 中國農業科學院深圳農業基因組研究所
發表時間: 2023年9月
發表期刊: Plant Communications
影響因子: 10.5/Q1
研究背景
景天酸代謝(CAM)具有很高的水分利用效率,被廣泛認為是從 C3 光合作用前進演化而來。不同的植物系趨同地前進演化出了 CAM,但 C3 到 CAM 前進演化的分子機制仍有待明確。鹿角蕨為研究從 C3 光合作用向 CAM 光合作用過渡的分子變化提供了一個機會,因為該物種同時存在兩種光合作用模式, 孢子葉(SL)和蓋子葉(CL)分別進行 C3 和弱 CAM 光合作用。
研究結果
1.鹿角蕨弱 CAM 的生理和生化復雜性
作者透過比較二歧鹿角蕨的孢子葉(SL)和蓋子葉(CL),以及典型的CAM和C3植物的氣孔導度、凈光合作用速率、蘋果酸含量和δ13C值等重要光合作用指標(圖1),發現weak CAM和strong CAM的氣孔導度和凈光合速率晝夜變化表現一致,但它們的蘋果酸晝夜積累模式和δ13C值存在明顯差異,這表明weak CAM途徑光合作用的復雜性,同時也暗示了weak CAM在自然界的存在可能遠比我們想象的普遍。
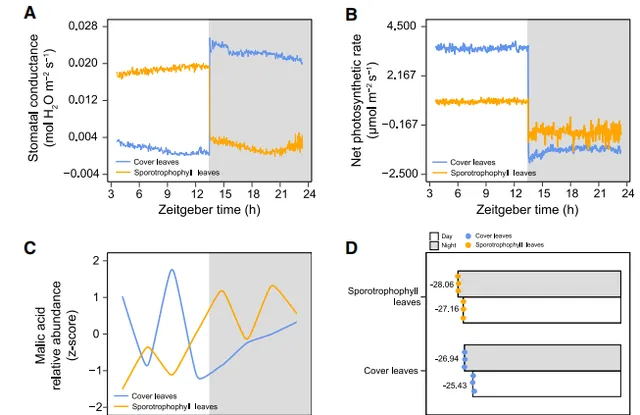
圖1 二歧鹿角蕨的孢子葉(SL)和蓋子葉(CL)光合特性
2.孢子葉(SL)和蓋子葉(CL)多組學變化
作者在12小時/12小時的光/暗周期中培養植物,每3小時采集1次孢子葉(SL)和蓋子葉(CL) ,進行轉錄組,蛋白質組,代謝組檢測。來研究CAM和C3葉片之間的分子差異。GC-MS檢出139中代謝物(圖2A-C)。 聚類熱圖,PCA分析顯示,相對於晝夜模式 ,初級代謝物在組織中顯示差異性更大(圖2B)。73個差異代謝物中,59個(80.8%)具有組織特異性效應,29個(39.7%)具有晝夜效應,27個(37.0%)存在組織特異性與晝夜效應的互動作用(圖2D,E)。隨後進行了轉錄組,蛋白質組分析,顯示CL樣本在PCA上有分離,說明CL樣本在這兩個層面上具有更強的時間調控性。
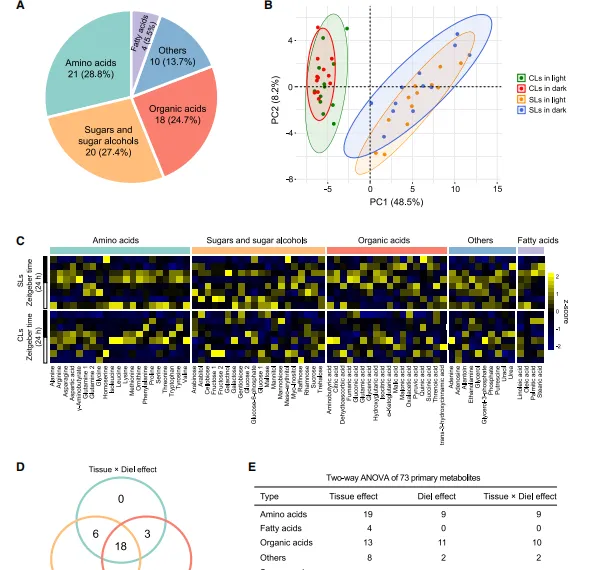
圖2 組織模式和時間效應對二歧鹿角蕨的影響
3.孢子葉(SL)和蓋子葉(CL)表現出不同的TCA迴圈晝夜動態
使用JTK_CYCLE方法研究了在這兩種葉片類別中特定表現出晝夜迴圈節律的初級代謝物。73種代謝物,CLs中有19種表現節律性,SLs中只有5種。與C3葉片相比,CAM葉片CLs中有更多的代謝物受到時間調控。在CLs中,三羧酸(TCA)迴圈中的三種有機酸被檢測為節律性代謝物:草乙酸、a-酮戊二酸和異檸檬酸。在SLs中,三種參與TCA迴圈的有機酸被鑒定為節律性代謝物:異檸檬酸、檸檬酸和琥珀酸。參與TCA迴圈的關鍵代謝物異檸檬酸在Cls和SLs中均顯示出節律性(圖3)。
在 代謝組 水平上,CLs 途徑中所有七種定量代謝物的濃度都在光照期間達到峰值。相比之下,SLs 中所有七種代謝物的濃度都在黑暗期達到峰值。在 蛋白質組 水平上,與TCA迴圈相關的蛋白質,包括檸檬酸合成酶、烏頭酸水解酶、異檸檬酸去氫酶、a-酮戊二酸去氫酶、琥珀酸去氫酶、富馬酸水解酶和蘋果酸去氫酶的豐度在CLs的光照期或黎明期達到峰值,但在SLs的黑暗期達到峰值。參與 TCA 迴圈的基因的晝夜表達譜與代謝組和蛋白質組數據的結果一致。綜合來看, 多組學數據表明 CAM 植物葉片在光照期間的 TCA 迴圈往往比在黑暗中更活躍。
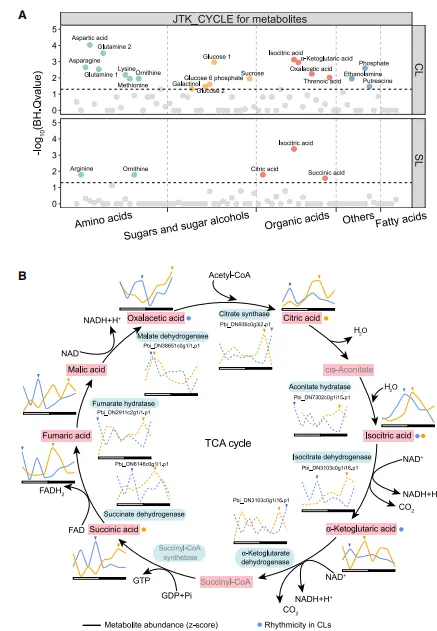
圖3 孢子葉(SL)和蓋子葉(CL)中TCA迴圈的變化規律
4.二歧鹿角蕨CAM通路的動態及轉錄調控
作者研究了CLs中與CAM途徑相關的基因表現和蛋白質豐度的晝夜動態。在 蛋白質組 中發現的所有七種核心CAM途徑蛋白質在 CLs 和 SLs 中的晝夜動態均呈反向模式(圖4A)。在具有 CAM 效能的CLs中,與夜間CO2同化和蘋果酸產生有關的蛋白質的表達在黑暗中達到峰值。相比之下,參與蘋果酸脫羧和 PEP 再生的蛋白質在光照下達到峰值,為卡爾文-本森迴圈產生CO2,並為重新進入CAM途徑提供受質。與CAM 途徑相關的基因轉錄組動態反映了蛋白質組分析。
二歧鹿角蕨中發現801個轉錄因子,在相同的基因組背景下,CAM途徑中的基因在具有CAM表現的CLs和具有C3表現的SLs中表現出相反的晝夜表達模式,這表明基因表現的轉錄調控可能是CAM途徑的關鍵因素之一。使用時間有序基因共表達網絡(TO-GCN)方法,以C3表現型SLs為對照,構建了CAM途徑的轉錄調控網絡(TRN)。透過選擇''C1+C2-'' GCN 集,即在 CLs 中具有正皮爾遜相關性而在 SLs 中具有負相關性的 TF基因對的重疊,確定了 786 個 TF、12 494 個蛋白編碼基因和 36 155個TF-基因對。根據這些 "節點 "和 "邊緣",提取 CAM 通路中的蛋白編碼基因及其共表達的 TFs 構建了假定的 TRN,確定了 39 個 TFs 作為調控二歧鹿角蕨CAM通路的候選基因(圖4B)。其中,bHLH TF 家族的基因數量最多,這意味著它們對 CAM 通路的調控具有重要意義。在 TRN 中還發現了5個ERF、2 個MYB、2個ARF、1個Dof 和1個WRKY,這表明CAM通路的轉錄調控是一個由多個TF 家族協調的精細而復雜的生物過程。
圖4 與CAM通路相關的蛋白質表達模式和轉錄調控網絡
5.CLs氣孔導度的時間動態和轉錄調控
CO2是與離層素(ABA)訊號通路共同調控氣孔運動的關鍵因子。PP2A和PP2C也透過調節OPEN STOMATA 1 (OST1)參與ABA感知復合物PYR1/PYL1/RCAR。作者發現,PP2A和PP2C在CLs和SLs之間呈現相反的表達模式。同時,在暗周期中,OST1的表達在CLs中增加直到21小時,而在SLs中保持不變。
氣孔運動也由保衛細胞的膨脹變化驅動。呼吸爆發氧化酶同源物(RBOHD和RBOHF)在CLs中在黃昏時達到峰值,在夜間保持低表達,但在SLs中的表達模式相反,表明它們可能在調控氣孔運動中發揮重要作用。這些結果表明,二歧鹿角蕨弱CAM葉片的倒掛晝夜模式是由與植物激素(如ABA)一致的離子和CO2濃度的守衛細胞感應觸發的,這一假設有待於透過生理實驗和對其他CAM物種的全面研究來進一步驗證。
用TO-GCN構建的TRN顯示,來自17個基因家族的26個TFs可能參與調控與CLs氣孔運動相關的11個蛋白編碼基因(圖5B)。有趣的是,來自 bHLH 家族的 TFs 數量最多(26個),這與CAM通路中TFs的家族分布一致。這表明,一些調控 C3 植物氣孔發育的 bHLHs 可能參與調控 CAM 物種的氣孔運動。然而,在兩個 bHLH TF 組之間沒有發現重疊。相反,四個 TF,即 MYB(Pbi_DN37017c0g1)、ERF(Pbi_DN6647c0g1)、ARF(Pbi_DN450c1g1)和 Dof(Pbi_DN294c1g1)被確定為同時調控 CL 中氣孔運動和 CAM 通路的潛在候選 TF。這些結果表明,二歧鹿角蕨的 CAM 通路與氣孔運動之間可能存在部份轉錄調控重疊。
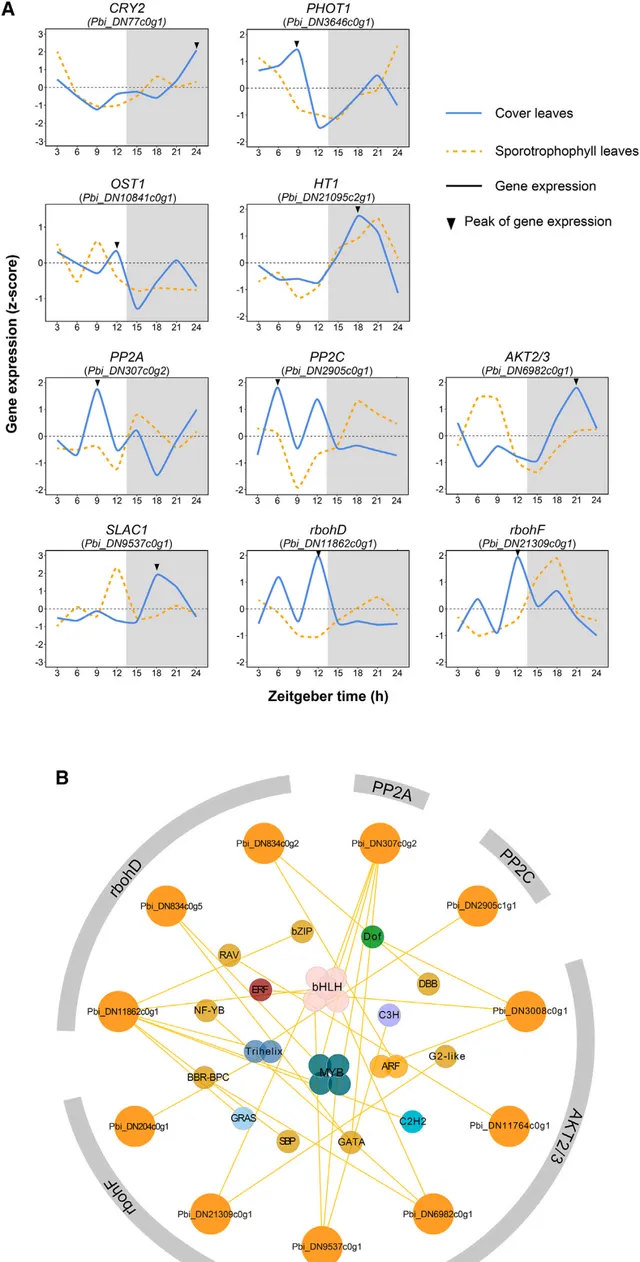
圖5 二歧鹿角蕨氣孔運動相關的基因表現模式和轉錄調控網絡
6.CAM植物中PPCK介導的PEPC磷酸化的表達趨同
CAM光合作用區別於C3光合作用的一個獨特特征是夜間二氧化碳同化。在CAM生理過程中,有四種蛋白質參與二氧化碳同化的酶促反應:b-CA、PPCK、PEPC 和 MDH。這四個基因的多個拷貝在黑夜周期中的表達量均高於白天。我們評估了這些基因在 CAM 物種間可能的趨同前進演化。b-CA 、PPCK、PEPC 和 MDH 基因的蛋白質序列在多個 CAM 物種之間沒有發現趨同現象。 在所有 CAM 物種中,PEPC 的序列趨同並非遵循相同的路徑。
整合了多個CAM物種中四個與CO2同化相關基因的已發表時間基因表現數據,以檢驗參與夜間CO2同化的基因在表達水平上是否經歷了趨同前進演化。PPCK的表達水平在白天相對較低,在黑暗時期逐漸升高(圖6)。PPCK在黑暗中使PEPC磷酸化,減少蘋果酸介導的對PEPC活性的抑制,從而增加夜間的二氧化碳同化。這些結果表明, PPCK對PEPC的磷酸化可能是CAM植物夜間CO2同化趨同前進演化的關鍵步驟,凸顯了基因表現調控在CAM途徑前進演化中的重要意義。
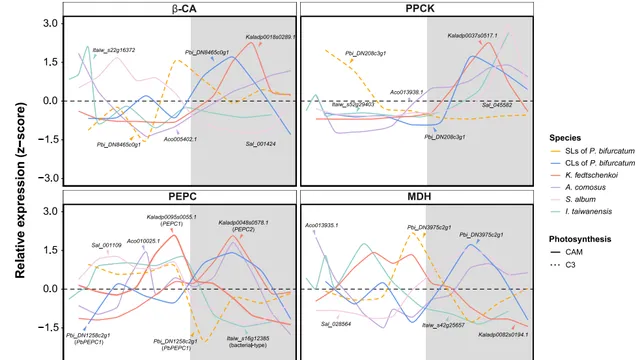
圖6 二氧化碳同化相關基因的表達趨同
邁維小結
在相同的遺傳背景和環境條件下,作者研究了 分別進行 C3 和弱 CAM 光合作用 的二歧鹿角蕨 孢子葉(SL)和蓋子葉(CL) 雙態葉片的代謝組、蛋白質組和轉錄組的晝夜動態。結果顯示。多組學數據表現出組織和晝夜效應。分析表明,與Sls相比,Cls中與能量產生途徑(TCA 迴圈)、CAM 途徑和氣孔運動相關的生物化學在時間上重新布線。還證實了磷酸烯醇式丙酮酸羧化酶激酶(PPCK)在高度不同的CAM系中表現出基因表現的趨同性。基因調控網絡分析確定了調控 CAM 通路和氣孔運動的候選轉錄因子。總之,研究結果為弱CAM光合作用提供了新的見解,並為CAM生物工程提供了新的途徑
往期熱門分享














