編譯:微科盟 雨,編輯:微科盟 景行、江舜堯。
微科盟原創微文,歡迎轉發轉載,轉載請註明來源於【轉錄組】公眾號。
導讀
幹旱脅迫嚴重制約辣椒( Capsicum spp.)的生長、產量和品質。獨腳金內酯(SL)是一種相對較新的植物激素,有望緩解辣椒植株的幹旱相關癥狀。然而, 關於葉面施用SL後暴露於幹旱脅迫(DS)時如何影響辣椒的基因表現的知識有限。為了探索這一點,本研究進行了生理學和轉錄組分析,以揭示SL減輕DS對辣椒幼苗影響的機制。結果發現,DS抑制辣椒幼苗生長,改變抗氧化酶活性,降低相對含水量(RWC),並引起氧化損傷 。相反, SL的施用顯著提高了RWC,促進了根系形態,增加了葉片色素含量。SL還透過降低MDA和pO2水平以及維持POD、CAT和SOD活性來保護辣椒幼苗免受幹旱誘導的氧化損傷 。此外, 轉錄組學分析顯示,在DS和DS+SL處理中,差異表達基因在核糖體、ABC轉運蛋白、苯丙烷生物合成和生長素/MAPK訊號通路中富集 。此外,qRT-PCR結果顯示,與僅幹旱處理相比,SL處理中 AGR7 、 ABI5 、 BRI1 和 PDR4 上調, SAPK6 、 NTF4 、 PYL6 和 GPX4 下調。特別是SL訊息傳遞的關鍵基因 SMXL6 在幹旱條件下被下調。這些結果闡明了SL介導的植物DS耐受性的分子方面,並為有效實作辣椒抗旱性提供了關鍵策略。
論文ID
原名: Physiological and Transcriptome Analysis of the Effects of Exogenous Strigolactones on Drought Responses of Pepper Seedlings
譯名: 外源獨腳金內酯對辣椒幼苗響應幹旱脅迫的生理和轉錄組分析
期刊: Antioxidants
IF: 7
發表時間: 2023年11月
通訊作者: 汪誌偉
通訊作者單位: 海南大學
DOI號 : 10.3390/antiox12122019
實驗設計
結果
1 SL緩解了幹旱脅迫對辣椒形態的負面影響
為了證實SL在DS環境中對辣椒生長的可能影響,研究人員檢測了對照組、DS組和DS+SL組的辣椒形態。 DS對辣椒幼苗根系生長有明顯的抑制作用。 SL預處理能有效緩解DS的抑制作用,在DS條件下,植株的根長、表面積、體積、根尖、根交叉和根分叉分別比對照處理減少49.56%、59.45%、65.27%、32.31%、55.7%和56.62%。 此外,SL預處理後,與對照組相比,減少率分別僅為20.13%、35.48%、35.71%、15.4%、29.11%和22.63%。重要的是, SL的葉面施用顯著促進了DS環境中辣椒幼苗的根系結構系統 (圖1)。 這些結果表明SL可以維持根系的正常生長和平衡吸水。
圖1. 辣椒幼苗的表型變化。 (A)辣椒植株和根系在不同處理下的表型。(B)根的總長度、(C)根表面積、(D)根體積、(E)根尖、(F)根交叉和(G)根分叉。對照:在正常對照條件下,DS:幹旱條件,DS+SL:施用SL的幹旱條件。數據是平均值和標準誤差;黑點表示三個生物復制品。不同的字母表示不同處理之間的顯著差異( p <0.05)。
2 葉片組織的顯微結構
研究人員評估了不同處理的植物的氣孔特性,以檢測SL對其在DS下氣孔開啟和關閉能力的影響 (圖2)。SEM顯示,與對照組相比,在僅幹旱處理中,植物氣孔完全閉合(圖2B)。在SL和DS條件下施用時,氣孔關閉減輕,表明外源SL可以改變DS引起的氣孔開放(圖2C)。此外,與對照組相比,氣孔面積減少了98%;然而,在幹旱脅迫下,SL葉面施用使氣孔面積增加了57.45%(圖2D)。 SL葉面施用可以透過調節細胞通透性來維持葉片中的水分水平,從而促進氣孔的開啟 。
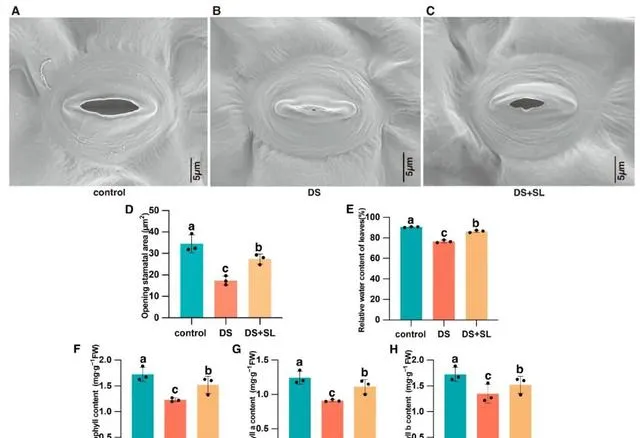
圖2. 在對照條件、DS(幹旱條件)和DS+SL(幹旱條件下施用SL)下,辣椒植株氣孔開度效能和生理分析的綜合效應。 (A-C)氣孔孔徑效能;(D)不同處理組氣孔面積的分析;(E)測定的相對含水量;(F)葉片的總葉綠素;(G)葉片葉綠素a;和(H)葉片的葉綠素b。數據是平均值和標準誤差;黑點表示三個生物復制品。不同的字母表示不同處理之間的顯著差異( p <0.05)。
3 葉片的相對含水量和葉綠素含量
幹旱嚴重影響了辣椒幼苗的RWC和葉綠素含量。 只有DS處理的RWC(76.57%)低於對照組(90.61%)。值得註意的是,用DS+SL處理的辣椒的葉片RWC(86.35%)顯著高於僅在幹旱條件下生長的辣椒(圖2E)。幹旱處理顯著降低了辣椒幼苗葉片中的葉綠素含量。與對照相比,辣椒幼苗中總葉綠素、葉綠素 a 和葉綠素 b 的濃度分別下降了28.8%、26.82%和21.91%。然而,與DS處理植物相比,DS+SL植物表現出相當高的葉綠素含量,分別增加了23.69%、22.33%和12.78%(圖2F-H)。
4 抗氧化酶活性和相關代謝產物的測量
DS +SL處理顯著影響辣椒幼苗抗氧化酶(POD、CAT和SOD)活性 。與對照組相比,DS顯著提高了POD、CAT和SOD活性。然而,預處理SL抑制了辣椒幼苗葉片中的酶活性。例如,DS的POD、CAT和SOD活性僅比對照組高180.86%、93.59%和107.02%。相反,DS+SL組的酶活性低於僅DS組(圖3A-C)。與對照組相比,僅DS組的脯胺酸、MDA和pO2顯著增加。相反,與DS組相比,DS+SL植物的脯胺酸、MDA和pO2含量顯著降低,分別為88.39%、57.89%和61.83%(圖3D-F)。在DS條件下,可溶性糖含量增加。此外,DS和DS+SL處理沒有顯著差異(圖3G)。
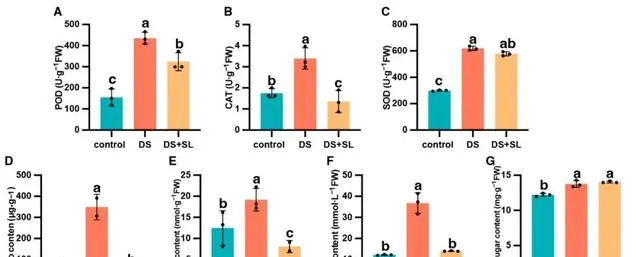
圖3. 對照、DS和DS+SL辣椒植株葉片中酶活性和非酶抗氧化劑含量的影響。 (A)POD、(B)CAT和(C)SOD的抗氧化酶活性;(D) 脯胺酸含量、(E)MDA含量、(F)pO2含量和(G)可溶性糖含量。數據是平均值和標準誤差;黑點表示3個生物重復。不同的字母表示不同處理之間的顯著差異( p <0.05)。
5 RNA-seq數據評估和差異表達基因分析
本研究的三組轉錄組測序共產生40320 kb原始數據,研究者對其進行低質素reads過濾,得到3869萬個clean reads。研究發現,88.58-93.36%的reads可以成功比對到辣椒( C.chinense )參考基因組,唯一比對率在85.35-89.02%之間(表S2)。層次聚類分析顯示,同一處理的不同樣本聚集在一起,DS+SL組表現出最顯著的差異(圖4A)。 轉錄組被分為四個DEG簇,每個簇中的基因具有相似的表達模式,分別包括95、1481、159和25個DEG (圖4B)。 在DS vs 對照組、DS+SL vs 對照組和DS vs DS+SL組中共鑒定出2097個DEG 。韋恩圖顯示,只有2個基因在所有組別中都有表達,而且在所有三個組別中,特異性DEGs都高於共享DEGs(圖4C)。此外,與對照組相比,DS組和DS+SL組分別有410個和849個DEG。此外,DS組和DS+SL組之間存在838個DEG(圖4D)。
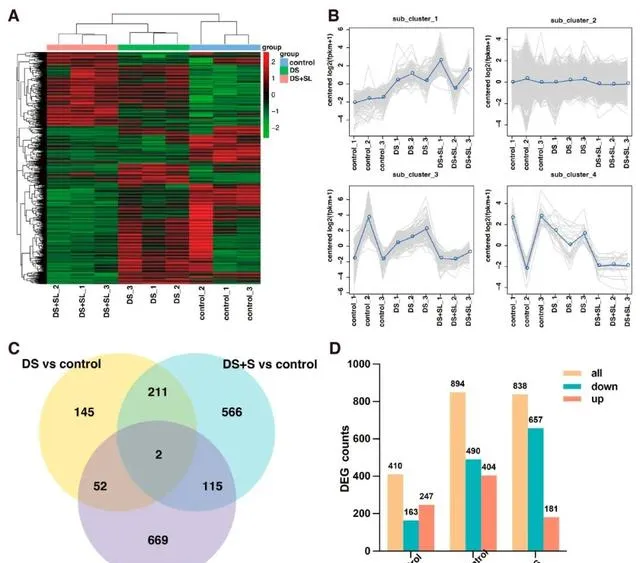
圖4. 三組在不同處理下的DEGs。 (A)DEG的熱圖和聚類分析表達,(B)RNA-seq數據的4個子聚類的放大區域。灰色線表示基因的相對校正基因表現水平,藍色線表示所有基因的相對糾正基因表現水平、(C)三組DEG的韋恩圖和(D)DEG的數量的一致性。
6 DEGs的富集分析
研究者比較對照組和DS組的DEG,發現後者富集在半乳糖代謝、植物激素訊息傳遞、光合作用和光合作用天線蛋白。當比較對照和DS+SL時,發現後者富集在胺基酸生物合成、油菜素甾體生物合成、類胡蘿蔔素生物合成和MAPK訊號傳導。 有趣的是,與對照組相比,DS和DS+SL組在核糖體途徑中表現出顯著富集。 DS和DS+SL處理之間鑒定的DEG富集在苯丙烷生物合成、萜類骨架生物合成、類黃酮生物合成、類胡蘿蔔素生物合成和MAPK訊號通路 (圖5、圖S1和表S3)。此外,如GO的分子和生物功能所描述的,感興趣的基因被分類為分子功能(MF)、細胞成分(CC)和生物過程(BP)。 研究者進行GO功能註釋,以充分了解DEG在對照組、DS組和DS+SL組中的作用 (圖S2)。
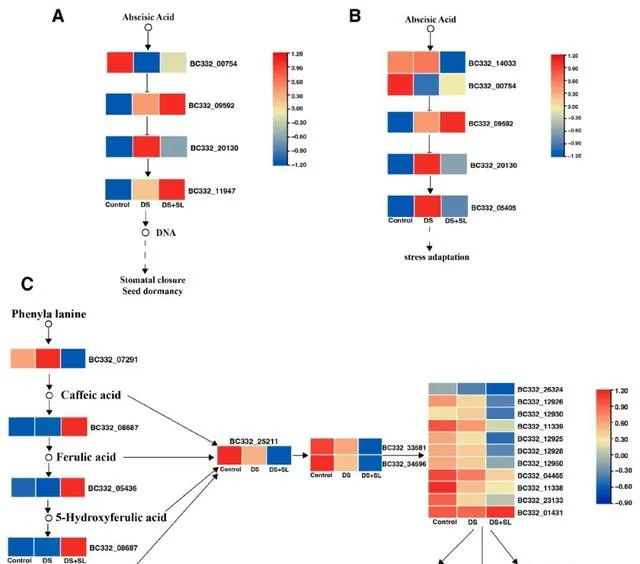
圖5. 與幹旱和SL處理相關的富集途徑。 (A) 植物激素轉導,(B)MAPK訊號,和(C)苯丙烷生物合成途徑。熱圖顏色從左到右顯示控制、DS和DS+SL的FPKM的log2值。
7 參與幹旱和SL處理的候選基因
基於富集分析,研究人員鑒定了71個基因,其表達在對照組、DS組和DS+SL組之間顯示出顯著差異,這表明它們可能對DS有反應或參與SL誘導的DS緩解。與苯丙類化合物生物合成途徑相關的基因在單獨的DS組和DS+SL組之間大量富集,並確定了18個差異表達基因。 有趣的是,與苯丙烷生物合成相關的基因 BC332_05436 ( cytochrome 84A1 )和 BC332_01431 ( peroxidase 15 )在DS+SL組中的表達顯著高於DS組植物。相反, BC332_25211 ( 4-coumarate -- CoA ligase 2 )、 BC332_33581 ( caffeoyl-CoA O-methyltransferase )和 BC332_34596 ( caffeoyl-CoA O-methyltransferase )在DS+SL中的表達顯著低於單獨的DS組。此外, 與對照組相比,在單獨DS和DS+SL處理的植物中,與植物激素訊息傳遞和MAPK途徑相關的基因顯著富集。 BC332_22503 ( indole-3-acetic acid-induced protein ARG7 )和 BC332_09592 ( BRI1 kinase inhibitor 1 )在DS+SL組中表達明顯,在DS組中表達較低。此外, 研究人員從DEG中鑒定了15個SL生物合成相關基因。參與SL生物合成的基因的表達譜在對照組、DS組和DS+SL組之間顯示出顯著差異。 與DS組相比, BC332_23620 ( SMAX1-LIKE 6 )、 BC332_15224 ( SMAX1_LIKE 7 )和 BC332_18735 ( SMAX1/LIKE 4 )在DS+SL中的表達更高。此外, 研究者在ABC轉運蛋白途徑中鑒定了23個基因 。與對照處理相比,基因 BC332_30010 、 BC332_33666 和 BC332_16040 在DS+SL處理中上調,但在DS處理的植物中下調(圖6)。此外,研究人員獲得了脯胺酸和抗氧化酶生物合成基因的mRNA水平,這在分子水平上進一步闡明了候選基因(圖S3)。
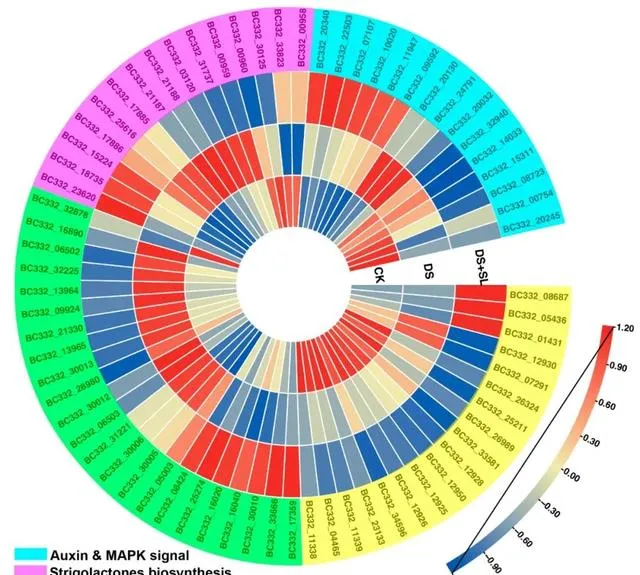
圖6. 與幹旱和SL處理相關的DEG分別在對照、DS和DS+SL中的表達。 熱圖顏色顯示FPKM的log2值。
8 基因表現的qRT-PCR驗證
研究者選擇15個與DS密切相關的候選基因用於透過RNA-Seq分析表達(表S4)。研究使用對照組作為對照,同時對DS組和DS+SL組進行研究,以驗證本研究中獲得的RNA-seq數據的可靠性(圖7)。
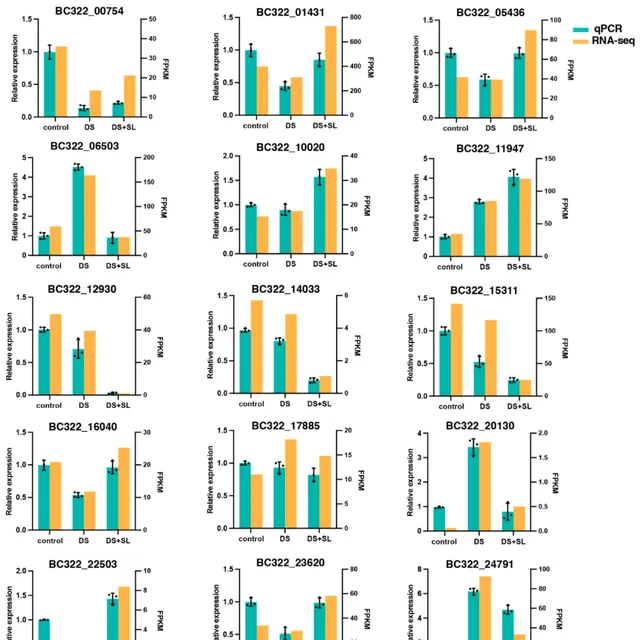
圖7. DEGs相對表達水平的qRT-PCR驗證。 數據是平均值和標準誤差;黑點表示3個生物重復。
討論
幹旱被公認為阻礙植物生長和發育的主要非生物脅迫因素。幾種植物激素在調節植物應激反應中的關鍵作用已得到廣泛認可。SL在調節植物發育和減輕環境壓力方面具有至關重要的作用。此外,SL的葉面施用透過改善光合特性和抗氧化酶活性來增強DS條件下的植物生長。然而,關於其對辣椒非生物脅迫反應的影響的資訊很少。
DS顯著抑制植物生長。 在本研究中,在DS條件下,辣椒幼苗的根和葉受到不利影響。SL預處理可以緩解辣椒的幹旱癥狀。在幹旱脅迫處理中,枯萎的幼葉表現出下垂和收縮的跡象,大多數成熟的葉片已開始卷曲,而DS+SL處理的葉片僅表現出輕微癥狀(圖1和圖2E)。 因此, 研究結果表明,SL的施用減少了DS條件下辣椒幼苗的明顯幹旱損傷。 植物中的葉綠素含量是光合活性的關鍵指標,但在脅迫條件下,這一特性會受到不利影響。DS顯著降低了葉綠素含量。此外,SL在調節植物光合效率方面發揮著至關重要的作用。 本研究結果表明,外源施用SL顯著緩解了DS條件下葉綠素含量的下降 (圖2F-H) 。因此,這表明SL是辣椒對抗DS的有效物質 ;在葡萄中也報道了類似的結果。 因此,施用SL很可能是一種可以用來減輕DS對辣椒幼苗不利影響的方法。
脯胺酸被認為是環境脅迫條件下植物細胞內合成的一種重要代謝產物。它被認為對在脅迫下生長的植物的保護功能有重大影響,這歸因於它作為滲透保護劑、膜穩定劑和ROS清除劑的能力。先前的文獻表明,脯胺酸積累可能有助於改善水分狀況,減少非生物脅迫引起的氧化損傷。同樣, 研究人員觀察到DS顯著增加了辣椒植株中脯胺酸的積累。相反,SL的施用顯著降低了DS條件下辣椒幼苗中的脯胺酸濃度 (圖3D), 這表明SL在DS期間對滲透調節具有潛在的效率效應,這可能有助於植物維持生長和功能 。脯胺酸生物合成基因( AtP5CS1 )在非生物脅迫條件下增加脯胺酸生物合成中起關鍵作用。研究人員發現,在有或沒有SL的幹旱條件下, P5CS ( BC332_15138 )的mRNA水平在辣椒中增加(圖S3)。 SL的套用導致脯胺酸含量顯著降低,這表明P5CS轉錄水平的上調可能有助於脯胺酸濃度的增加。 然而,SL影響脯胺酸的機制尚不清楚,需要進一步探索。
在受到環境脅迫的植物中,維持氧化還原穩態對於減輕ROS的過度產生和最大限度地減少細胞膜損傷至關重要。 在本研究中,DS下pO2和MDA含量顯著增加,而SL透過抑制pO2和丙二醛含量的積累來逆轉DS誘導的辣椒幼苗的氧化損傷 (圖3E,F)。SL介導的幹旱反應可能涉及pO2含量和氣孔關閉的復雜相互作用。植物可能有必要維持某些基因的表達,以將氣孔開放度維持在特定水平,從而在缺水條件下平衡CO2流入和水分損失。研究人員認為,在DS過程中,氣孔被牢牢關閉。然而,SL的套用在緩解氣孔關閉方面有顯著效果(圖2C),表明SL在調節植物的水分平衡方面發揮著至關重要的作用,導致氣孔在DS下重新開放。這是一個復雜的分子機制,仍需進一步研究。為了應對氧化損傷,植物已經開發出一種成熟的抗氧化防禦機制。研究表明, SL可以增強抗氧化酶對ROS解毒的活性,從而賦予作物耐受性。與DS組相比,在DS+SL組中檢測到較低水平的POD、CAT和SOD活性,這表明SL可以提高ROS清除效率,並可能保護植物免受DS的不利影響,從而減輕DS引起的氧化應激(圖3A-C)。 此外, 研究人員評估了過氧化物酶4( POD4 )和過氧化物酶( POD15 )的表達水平,它們是參與苯基丙烷生物合成途徑的基因。 在幹旱暴露的幼苗中, POD15 的表達降低,而SL的施用顯著增加了這些基因的表達水平,這與轉錄組學結果一致。相反,SL處理後, POD4 幾乎沒有表達(圖7)。 POD 基因的轉錄水平是否有助於提高抗旱性或阻礙抗旱性值得進一步研究。
SL有效調節植物生長,以應對各種環境條件。阿拉伯芥的生理和分子分析表明,植物激素訊號通路對調節幹旱或缺水反應至關重要。 OSRK1 ( SAPK6 )是水稻根系脅迫訊號傳導的上遊調控因子,在ABA和高滲脅迫訊號傳導中發揮重要作用。 本研究發現,用SL預處理的植物在DS過程中表現出與植物激素訊息傳遞和MAPK訊息傳遞途徑相關的顯著基因變化 (圖5和圖6)。具體而言,研究人員觀察到Aux/IAA家族生長素反應蛋白的表達變化,這些蛋白在植物脅迫反應中起著關鍵作用。ROS濃度的增加對IAA turnover周轉產生負調控影響。有趣的是, 在本研究中,研究者發現DS組 ARG7 ( BC332_22503 )的表達水平降低了97.9%,而DS+SL組則顯著升高 。此外,發現 BC332_20130 ( SAPK6 )編碼激酶在沒有SL處理的DS反應中上調,但在SL預處理的組中顯著下調。研究人員假設這些基因可能透過SL的作用來減輕DS。
ABC轉運蛋白現在被認為參與了許多生理過程,使植物能夠適應環境變化。多能耐藥(PDR)亞家族ABC蛋白存在於許多植物中,可應對生物和非生物脅迫。先前的一項研究表明,ABC轉運蛋白 AtABCG36/AtPDR8 的過表達使植物比野生型植物更能抵抗幹旱和鹽脅迫。相反,敲除系比野生型植物對DS更敏感。 研究人員發現,在DS條件下SL預處理後, BC332_16040 ( PDR4 )顯著上調,這意味著 PDR4 可能在對DS和SL的反應中發揮關鍵作用。 先前的研究表明, MAX2 和 DWARF14 之間的相互作用觸發SL的訊息傳遞,而 SMXL6 ( SUPPRESSOR OF MAX1-Like 6 )起到SL訊息傳遞抑制劑的作用。 MAX2 突變體對DS表現出高度敏感性。最近的一項研究發現,阿拉伯芥中 SsMAX2 的過表達顯著增強了對幹旱、滲透和鹽脅迫的抵抗力。此外, SMXL6 、 SMXL7 和 SMXL8 在阿拉伯芥DS過程中發揮負調控作用。 研究人員發現 BC332_17885 ( CcsMX1 )的表達水平在DS下上調,但在SL處理下下調(圖7),表明其在DS反應中的負調控作用 。相反, CcsMX26 的表達呈現相反的趨勢。對行動機制的調查是難以捉摸的,需要進一步調查。
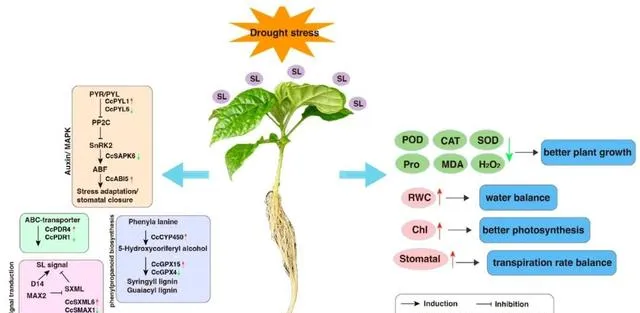
圖8. SL誘導辣椒幼苗耐受幹旱脅迫的機制綜述。
總之, 本研究的綜合研究,包括表型觀察、生理測量和基因表現分析,證明了SL對DS環境中辣椒植物的多方面積極影響。SL有效地減輕了DS的不利影響,並在這種具有挑戰性的條件下促進了辣椒幼苗的生長 。 SL的套用是對抗幹旱誘導的氧化損傷和增強對DS耐受性的有力工具 ,最終促進辣椒植物的整體健康發育。此外, 本研究結果強調,對DS反應至關重要的DEG主要與MAPK和植物激素訊息傳遞途徑有關 。這些見解有助於更深入地理解辣椒抵抗DS能力的分子和生理機制,並為闡明SL如何在辣椒對幹旱脅迫的適應力反應中與不同的訊號通路復雜相互作用提供了一個有價值的概念框架(圖8) 。這項研究為未來研究在面對環境挑戰時提高作物的復原力和可持續性奠定了堅實的基礎。











