編譯:微科盟未知,編輯:微科盟Emma、江舜堯。
微科盟原創微文,歡迎轉發轉載。
導讀
轉錄組、蛋白質組和代謝組構成了植物基因型穩定性的柔性物質基礎,使其能夠產生隨環境變化而動態變化的表型。將轉錄組、蛋白質組和代謝組嵌入植物代謝途徑通常需要進行復雜且異位的生物技術實驗。由質譜成像(mass spectrometry imaging,MSI)主導的空間多組學技術透過相關轉錄因子、功能性蛋白質和代謝物在通路中的空間關聯,提供了最直觀的原位證據鏈。這種空間證據可以與空間、時間、植物形態、組織器官相關聯,並為植物表型變化與其大分子或小分子組分之間的動態關系提供最直接的生理證據。本文介紹了MSI如何實作高通量原位空間代謝數據采集的儀器基礎以及影響成像質素的因素。接下來的內容涵蓋了過去3-4年的空間證據,總結了特定的植物分子生理套用,包括植物天然產物的空間生物合成途徑、植物根與外部環境之間的物質交換、植物在非生物脅迫下的生理抵抗、植物在生物脅迫下的化學防禦以及植物與微生物之間的相互作用。最後,討論了將空間多組學與表型技術結合,以實作未來植物生理研究中基因型-轉錄組-蛋白質組-代謝組-表型的時空一致性的可能性,並探討了從空間數據中挖掘植物生理資訊的困難。
論文ID
原名: Mass spectrometry imaging: an emerging technology in plant molecular physiology
譯名: 質譜成像:植物分子生理學的新興技術
期刊: Phytochemistry Reviews
IF: 7.7
發表時間: 2023.11
通訊作者: 李林楠,徐紅
通訊作者單位: 上海中醫藥大學中藥研究所
結果
1 介紹
為了探究植物中穩定基因型與動態表型之間的適應力代謝系統,將轉錄組、蛋白質組和代謝組相結合的多組學模型,已成為一種流行的技術。多組學提供的多個大型數據集可以從基元水平、方法學水平糊數學水平上整合和提取有效資訊。此外,多組學技術在探索植物在環境中動態表型與穩定基因型之間的調控方面取得了重大突破。然而,每種植物都具有動態的整體內部代謝系統,使其能夠靈活改變表型以維持穩態。地球上超過391000種植物產生數百萬種代謝產物。從轉錄組、蛋白質組和代謝組的大型數據庫的有效整合仍然是多組學發展的關鍵問題。近年來,一些微小分子的空間定位方法已經推動了與時間、空間和植物形態等維度相關的新內容的發展,以研究植物基因型和表型之間的物質代謝。
在植物生理學的發展中,視覺資訊通常被視為最直觀的補充證據。此外,自1920年代以來,對植物結構視覺化的探索方法一直深入到微觀水平(圖1a)。植物視覺化已經發展到可以在組織、器官、細胞,甚至亞細胞水平精確定位植物代謝途徑中的轉錄組、蛋白質組和代謝組的程度。質譜成像(MSI)起源於物理學和材料科學,透過開發能夠直接從樣本中提取分析物的離子源,如脫附電噴霧電離(desorption electricity spray ionization,DESI)和基質輔助激光解吸/電離(matrix-assisted laser desorption/ionization,MALDI)。然後,MSI套用於醫學和生物學領域,在顯微鏡和免疫組化技術的幫助下,逐漸成為生物組織中蛋白質和肽的空間分析的熱門方法。近年來,MSI已廣泛用於植物和天然產物的空間代謝組學。根據最近的文獻,生物組織的MSI主要用於藥代動力學,以研究藥物成分在實驗動物中的吸收、組織傳遞和代謝轉化的過程。這項研究的關鍵挑戰是排除來自動物內源成分的顯著幹擾。相比之下,植物MSI傾向於對植物代謝的組織進行全面的空間表征;在這裏,困難在於組織切片以及植物代謝的多樣性和復雜性。基於MSI,簡單處理的植物可用於收集化學成分的原位空間分布數據。透過質素對電荷比(m/z)鑒定的每個代謝物離子可以透過顏色差異的點陣矩陣來表征在組織中的豐度分布,從而可以同時實作多個小分子的組織定位(圖1b)。
與其他空間表征方法(如顯微檢查和免疫組織化學)相比, MSI具有許多優勢,包括無可比擬的便利性、高通量和廣泛適用性,可用於研究植物代謝多樣性。除了生物樣本中的蛋白質、脂質、核酸、醣類和激素外,堿性化合物、類黃酮、寡糖和植物激素的定性和定量分析也在基於MSI的空間代謝組學的套用範圍內。MSI不僅在原位分析方面具有廣泛適用性,而且在無需標記的情況下同時分析多個組分方面具有高通量和高效率的特點,這是其他原位分析技術所無法比擬的。近年來,基於MSI的優勢,植物學家一直致力於開發一種完整而高效的空間多組學研究模型,以探索植物在環境中動態表型與穩定基因型之間的調控。這一空間多組學模型還被套用於確定植物多種生理和生化狀態的微觀代謝密碼。植物中的代謝途徑可以在組織、細胞甚至亞細胞水平上還原為三維結構。
近年來,基於MSI引領的空間多組學研究模型揭示了植物在不同應激和短期或長期應對機制下的代謝調整,這些機制反映了植物與不同相關微生物之間的代謝通訊差異,以及植物對不同類別外源物質的代謝途徑差異。我們使用關鍵詞「質譜成像和植物成像」在Web of Science和PubMed數據庫中進行檢索,並在2019年至今檢索到60篇相關研究,本綜述介紹了MSI技術的基本原理和實驗程式,並總結了過去3年基於MSI的空間代謝組學在研究代謝物、植物生理和生物化學關系方面的主要套用,包括植物天然產物的空間生物合成途徑、植物根與外部環境之間的物質交換、植物在非生物脅迫下的生理抵抗、植物在生物脅迫下的化學防禦以及植物與微生物之間的相互作用。我們還從關鍵指標的角度討論影響影像質素的因素,並展望了基於MSI引領的空間多組學研究模型結合遙感等現代表型技術,在未來可能取得的對植物在環境中動態表型與穩定基因型之間調控的突破。
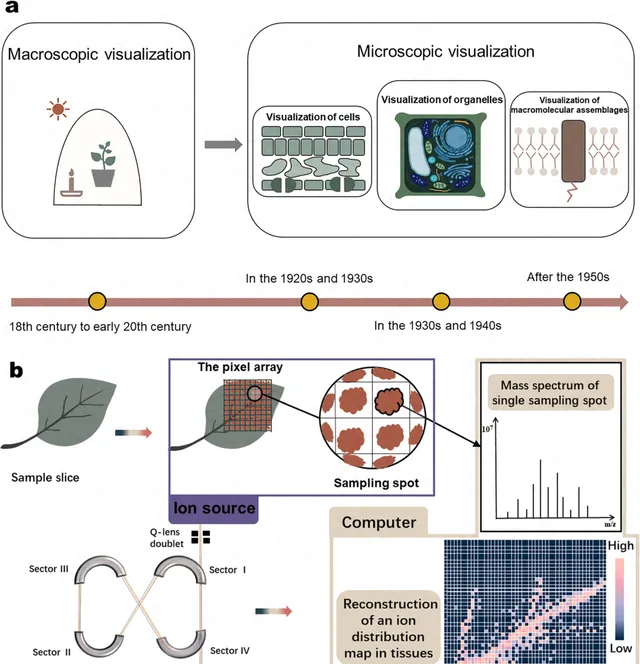
圖1 基於MSI的植物微觀視覺化和植物代謝物空間表征的歷史。(a)近100年植物微觀視覺化的發展(視覺化從宏觀到細胞、到亞細胞、到大分子的發育過程);(b)MSI重建代謝物分布圖的流程。
2 質譜成像技術的原理、實驗步驟和影響因素
2.1 MSI 原理和不同的電離類別
質譜成像儀主要由離子源、質譜分析器、探測器和數據處理系統組成。相較於其他質譜儀,質譜成像儀可利用特殊的離子源記錄待測離子的空間座標,並透過電腦的儲存、辨識、篩選和分類功能,重建離子在組織中的分布圖。除了常規參數如采集速度和質素分辨率外,還考慮了空間分辨率和成像的可重復性。MALDI、DESI和二次離子質譜(secondary ion mass spectrometry,SIMS)是分析植物小分子組分的三種最常用的離子源。表1提供了幾種離子源的具體參數。
表1三種常用離子源的比較

2.1.1 基質輔助激光解吸/電離 (MALDI)
由於離子源的電離過程,MALDI-MSI所使用的樣品在MALDI-MSI中有一個額外的步驟,即在樣品上噴塗基質。但總體方法相似。典型的MALDI-MSI工作流程如圖2b所示。透過適當的方法獲得的適當厚度的組織切片被放置在銦錫氧化物(ITO)塗層的載玻片上。選擇適當的MALDI基質,並均勻噴灑或昇華到組織表面,組織中的分析物最初在溶劑中溶解。隨著溶劑的蒸發,基質和分析物根據分子間力(包括氫鍵、鹵鍵、範德華力和π-π堆積)形成超分子結構的晶體。帶有共結晶樣品的載玻片被放入真空幹燥器中,以防止吸濕。然後,帶有共結晶樣品的載玻片被放入MALDI質譜儀進行分析。MALDI-MSI的電離過程發生在高真空環境中。首先形成基質離子,然後在激光束照射在玻片圓形區域上的情況下,吸收能量的分析物被汽化。在氣化過程中,代謝物透過基質離子與分析物之間的碰撞被離子化並進入四極桿分析器。透過掃描整個樣品,激光束可以跨越整個組織切片進行原位分析。在電離過程中,激光脫附具有強烈的電離效應,但來自組織的分析物可以透過基質離子的間接電離。相對較溫和的電離保持了電離效率,而空間分辨率主要取決於激光斑的大小,通常為5-10μm。大氣壓-基質輔助激光解吸/電離(AP-MALDI)是一種類似於MALDI的電離源;兩者都具有相同的樣品預處理步驟,並且都需要噴灑基質來協助分析物的電離。AP-MALDI與MALDI的區別在於,分析物的汽化和電離發生在大氣壓下。AP-MALDI是一種較為柔和的電離方法,更適用於不穩定化學成分,並且可以產生更穩定的離子流。AP-MALDI的大氣壓電離使基質離子能夠與分析物離子形成更均勻的電子雲,並且空氣對流起到冷卻效果,降低了由激光灼熱引起的分解損失。此外,AP-MALDI的靈敏度主要由大氣壓電離源和真空質譜分析器之間的離子傳輸效率決定,這通常需要選擇具有更大噴嘴和更高樣品消耗的AP-MALDI。
2.1.2 解吸電噴霧電離 (DESI)
DESI是一種電離源,其原理在理論上與電噴霧電離(ESI)相似(圖2a)。透過使用高電壓電場對溶劑進行帶電。使用噴霧氣體(N2)的氣動霧化器不斷向組織中噴灑溶劑液滴,導致其路徑中的電荷積累和溶劑蒸發。這導致了重復的庫侖爆炸和更小的帶電溶劑液滴。帶電的小溶劑液滴透過快速移動的霧化氣體(N2)引起的氣體噴射影響組織表面。帶電液滴在樣品表面的撞擊點溶解代謝物,然後碰撞、分解並噴射進入噴嘴口。在這些過程中,撞擊點處的分析物被電離,而溶劑繼續揮發,空間分辨率約為50μm。MSI的定位精度和可重復性受到溶劑毛細管的幾何特征(如剛度和尖端質素)的顯著影響。此外,透過最佳化溶劑組成、流速和幾何因素,還可以在不改變噴霧器結構的情況下在一定程度上提高DESI的空間分辨率和效能。
2.1.2 二次離子質譜 (SIMS)
SIMS中的電離是在組織表面發生的陰離子級聯碰撞過程(圖2c)。在高真空中,離子發射器將一束具有高動能的離子直接照射到樣品表面。在樣品表面的撞擊點上,被稱為電荷初級碰撞離子的表面粒子在電離過程中獲得動能,並行生異位碰撞產生二次碰撞離子。小的二次離子被濺射到註入器中,並在高速離子轟擊過程中到達質譜分析器。納米級二次離子質譜(Nano-SIMS)和飛行時間(TOF)-SIMS是兩種常見的質譜技術。透過在樣品表面進行高劑量離子轟擊,nano-SIMS可以達到的最高橫向空間分辨率為50納米。使用較低的離子劑量進行電離過程的TOF-SIMS無法達到這種高空間分辨率,但可以將對樣品表面的損傷降低到單個原子或分子層的1%。此外,TOF-SIMS適用於有機化合物和單原子離子的分析,因為既可以檢測低質素片段,也可以檢測相對較高質素的片段,顯示了廣泛的套用。TOF-SIMS還可以進行多組分-多系統的同時分析、深度剖析分析、3D成像等功能,使其在生物成像中得到更廣泛的套用。
2.2 植物樣品MSI的實驗程式
在通用的二維質譜成像技術中,透過簡單的預處理方法(如切片或壓印),盡可能在植物樣品中保留所有代謝物在同一平面上。載有該平面的載體被放入MSI儀器,以預設的空間分辨率進行數據采集。空間分辨率表示在同一平面上兩個像素可以被區分的最小距離,直接反映了儀器對空間區分的能力。空間分辨率通常透過座標方塊矩陣進行視覺化。每個座標點中的代謝物可以透過不同的離子源使用不同的電離方法進行收集,並最終進入質譜分析器以收集相關的質譜資訊。為了建立總離子流程圖,將每個像素中發現的所有片段離子及其獨特的電荷比率匯總。電腦軟件可以提取在誤差允許範圍內具有相同m/z值的每個離子的所有像素位置,並將它們儲存在每個離子座標數據集中,用於重建離子分布圖(圖1b)。
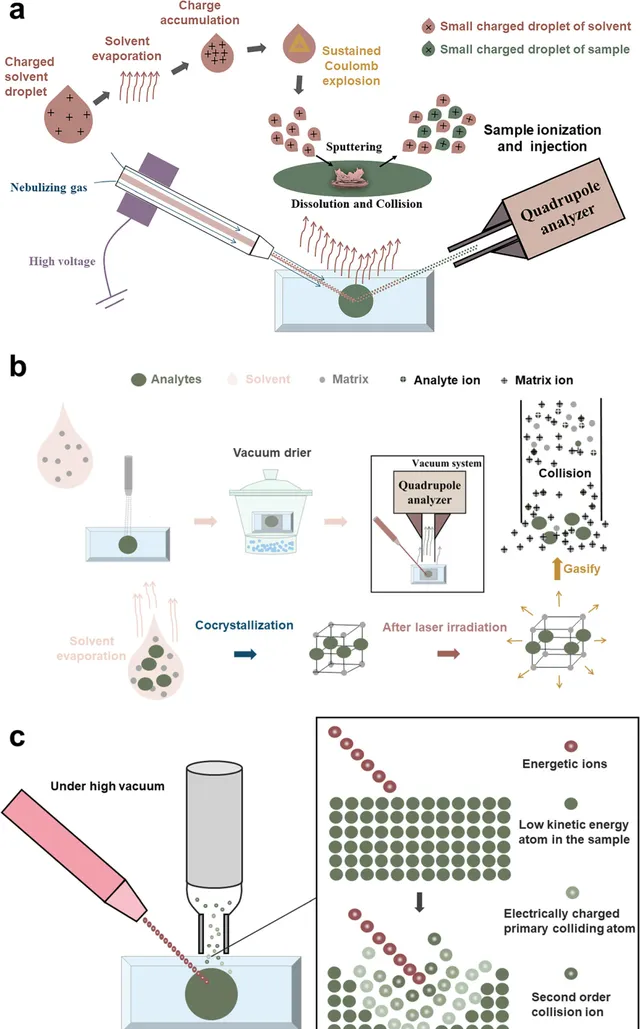
圖2 基於MSI的植物微觀視覺化和植物代謝物空間表征的歷史。(a)DESI原位電離示意圖(樣品表面分析物在濺射到進樣口之前在帶電微滴的作用下電離的過程);(b)MALDI的顯微演示(MALDI操作程式和基質輔助電離原理); (c)基於離子級聯碰撞的 SIMS(離子束轟擊下樣品表面分析物電離的微觀過程 )。
2.3 質譜成像的影響因素
MSI可以提供化合物的傳統質譜資訊和離子的空間座標資訊。基於期望獲取兩種相應數據的前提,我們需要從質譜和空間兩個維度的關鍵指標來理解影響MSI的因素。影響質譜成像影像質素的主要變量包括樣品完整性、分辨率、靈敏度和成像重復性。
2.3.1 樣品制備
對於許多使用冷凍微切片法作為預處理方法的樣品,獲取具有適當厚度、完整形態且無汙染的切片是成像質素的關鍵因素。切割的組織需要在適當的剛度狀態下準備,並且能夠被冷凍微切片機輕松切割成微米級的切片。有時,從生長中的植物中切取新鮮樣本,其含水量過多或組織不夠緊實,容易被冷凍微切片機的刀片碾碎。這些新鮮組織可以被冷凍在-80°C以稍微降低水分含量,並增加植物組織的緊密度,或者它們可以嵌入到適當的物質中。目前,最優切割溫度(optimal cutting temperature,OCT)化合物是最常見的嵌入介質之一。然而,如果組織完全嵌入到OCT中,它將成為導致離子抑制問題的汙染物,影響質譜分析。一些最近發現的嵌入介質可以在不影響質譜數據的情況下協助冷凍切片,包括羧甲基纖維素(carboxymethyl cellulose,CMC),這是纖維素和明膠、明膠、冰、蛋清或煮沸介質的混合物,提供了解決冷凍切片問題的潛力。此外,一些木材組織由於含水量低、纖維含量高、無黏性以及難以獲取新鮮樣本,過於堅硬難以切割成切片,需要新的技術或新的包埋介質。
2.3.2 空間分辨率
MSI中最關鍵的一個方面是其空間分辨率,這是理解MSI數據的一個重要概念。對於2D成像,需要考慮橫向空間分辨率;這個分辨率表示在同一平面上兩個像素可以被區分的最小距離,直接反映了儀器對空間區分的能力。儀器的空間分辨率是基於一般認識的,即樣品區域和兩個采樣點之間的最小距離都不會影響電腦能夠辨識的座標。通常報告的預設分辨率實際上是一個2D像素陣列,它實際上是樣品表面上采樣點的密度。MSI儀器在每個座標點獲取並記錄提取的分析物離子的質譜,包括提取分析物離子的所有座標點的位置,以便可以恢復原始組織中目標離子的分布(圖1b)。
基於樣品斑點大小和間隔距離這兩個主要因素,有許多技術用於提高MSI的空間分辨率。根據過去10年的研究,使用MSI離子源結合小孔、多焦距透鏡、光束擴充套件器或更改光束傳輸選項,已經可以定位個別細胞或亞細胞結構,甚至在三維中實作。這有可能取代費力的連續切片。使用自動聚焦光譜對不規則葉子樣本進行逐層采樣,建立亞微米深度空間分辨率,從而實作 3D MSI。根據最近文獻的檢索,3D質譜成像在植物生理學中並沒有廣泛套用。
這有潛力替代繁瑣的連續切片。Dreisbach等使用自動對焦光譜學逐層對不規則葉片樣本進行采樣,建立了亞微米深度的空間分辨率,從而實作了3D MSI。根據對近期文獻的搜尋,3D質譜成像在植物生理學中並沒有廣泛套用。
2.3.3 質素分辨率和質素靈敏度
質譜儀在兩個離子之間能夠分辨的最小m/z差異反映了儀器的質素分辨率;這個分辨率是影響成像離子通量和質素準確性的最關鍵因素。相等大小的兩個峰被認為在半峰的高度處分開,而在半峰的高度處的峰寬是質素分辨率的數學表示。質素分辨率的套用意義主要體現在質譜儀對不同m/z離子的分辨能力、辨識離子的m/z範圍的大小以及待測離子的質素準確性。這些對於不同類別的質譜成像分析儀器的質素至關重要。以下討論了幾種常見的質素分析儀器,表2提供了幾種質譜分析儀器的具體參數。
表2 不同質素分析儀的效能(質素分辨能力和質素精度主要針對m/z>1000)
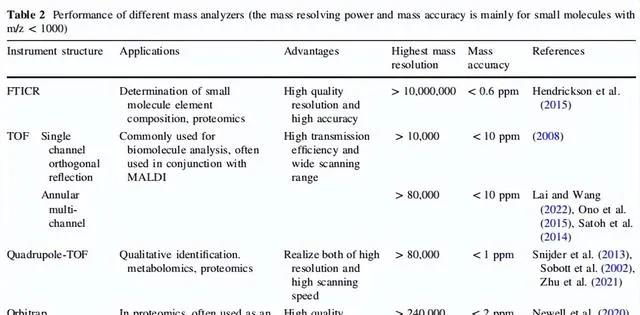
在相同的初始位置經過脈沖直流電場後,待測離子具有相同的初始速度並在飛行管中自由移動;然而,由於其不同的m/z值,離子到達探測器的時間是不同的,基於此概念TOF不斷得到改進。由於正交速度可以使待測離子在垂直於原始行進速度的方向上具有相同的初始速度。在離子在第一個垂直方向的初始速度降為零後,由於另一個電場,離子的速度調整到相反的方向,然後離子在第二個飛行管中自由移動以到達探測器。這個過程基本上使離子的飛行長度增加了一倍,而不會增加儀器的尺寸。在以上原理的支持下,單鍊結正交反射飛行時間質譜儀的質素分辨能力可以超過10000。隨著時間的推移,由於環形多通道TOF-質譜儀的出現,這種類別的質譜儀的質素分辨能力已經提高到了超過80000;然而,由於通道的延伸,m/z檢測範圍也相應減少了。目前,TOF經常與自身或其他質素分析儀器串聯以實作更高的空間分辨率、更寬的m/z檢測範圍、更準確的質素檢測準確度和更靈活的工作模式。
Orbitrap也是一種高分辨率質譜儀。這種質譜儀靈感來自Kingdon陷阱並可視為其升級版本。在20世紀90年代,這種基於靜電場的軌跡陷阱被認為對於質譜分析是有用的。兩個軸對稱電極巢狀在一起形成一個空間電場,外部電極為正,中心電極為負。透過精確設計這兩個巢狀電極的形狀,待測離子在電場的作用下以垂直於電場線的角度進入,離子同時進行穩定的軸向旋轉和徑向振蕩。因此,圍繞中心電極形成了穩定的離子軌域。精確的離子m/z可以從與初始離子速度和位置無關的軸向旋轉頻率中得出。基於這個Orbitrap,可以實作高質素分辨率。目前,Orbitrap在m/z 200處可以提供高質素分辨率(>240000),質素準確度<2 ppm;然而,其相對較慢的數據采集速度是一個限制。OrbiSIMS是一種新型質譜儀,它結合了TOF的快速采集速度和Orbitrap的高質素分辨率,突破了兩者的限制。
目前,傅立葉變換離子回旋共振(Fourier transform ion cyclotron resonance,FTICR)質譜儀提供了最高質素分辨率。這基於運動點電荷在均勻電磁場中的電磁場理論。待測離子在均勻磁場中加速其回旋運動,透過時變電場的作用被激發到更大的軌域,然後被檢測到。基於質譜儀原理的緊密擬合,透過共振修正了離子回旋運動的軸線,產生了可檢測的電訊號的空間幹涉。旋轉的離子在極窄的旋轉半徑內可以獲得非常大的平移能量,這是FTICR超高質素分辨率的關鍵。
總之,離子采集效率、空間分辨率、成像重復性、質素分辨率和質素靈敏度等參數需要被用作關鍵影響指標。在大多數情況下,速度和靈敏度不能同時達到最佳狀態;因此,需要根據實驗需求選擇速度或準確性,另一個則需要被犧牲。樣品采集速度主要由像素點面積、像素點之間的距離和每個像素點的采集時間決定。透過減少像素的數量(增加像素點的面積或增加它們之間的距離)或減少采集時間,可以獲得更高的采集速度。前者將導致較低的空間分辨率,而後者將要求質譜儀具有更高的靈敏度。對微代謝物的高靈敏檢測主要與離子源的電離效率、離子傳輸效率和質譜儀的離子檢測有關。質素分辨率主要與離子源和質素分析器中的精密結構有關,影響電離/解吸和質素分辨率能力。
3 MSI在植物分子生理學中的套用
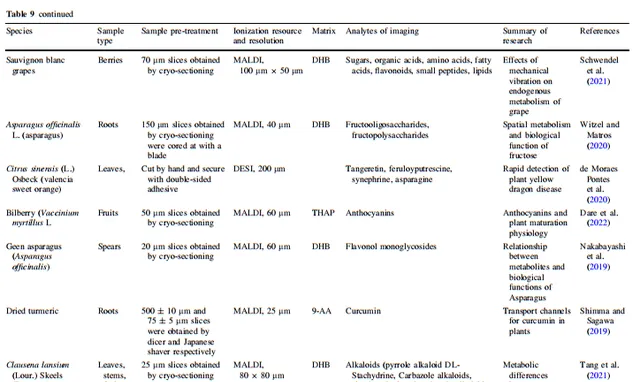
高等植物的基因組、轉錄組、蛋白質組和代謝組構成了彈性調控網絡,負責維持它們的正常生命過程。除了利用代謝物的響應位置來確定生物合成途徑中前體化合物、中間產物和產物之間的關系外,植物質譜成像(MSI)的空間代謝資訊還能反映植物在內源和外源化合物方面的實際執行狀態。透過分析植物組織中代謝物的聚集趨勢,可以辨識與其相關調控途徑中連線的功能性物質。在本節中,我們詳細總結了MSI在分析植物在非生物和生物脅迫下的脅迫耐受性和化學防禦、植物與外界物質交換、植物與微生物相互作用、植物內源代謝物生物合成途徑與生物學功能之間的關系、植物與微生物相互作用、植物殺蟲劑代謝以及空間生理資訊等套用方面的質素。所有植物質譜成像套用範例中涉及的離子源、空間分辨率、樣品處理方法和成像分析物類別在表3、4、5、6、7、8和9中都有詳細總結。
表3 植物在非生物脅迫下的耐逆生理
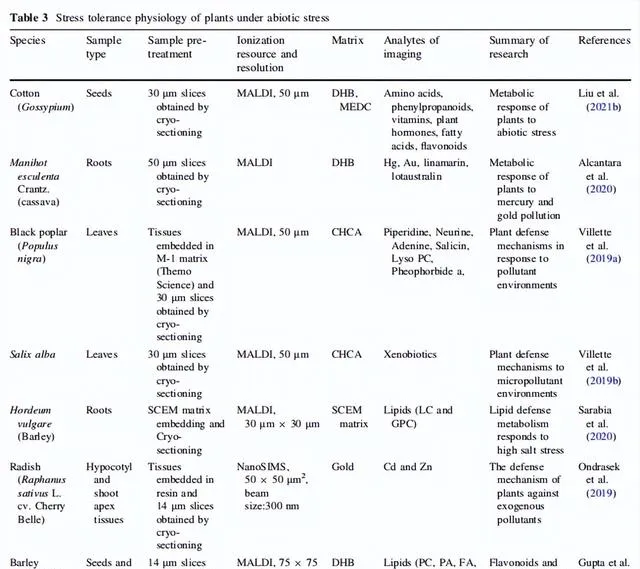
表4 生物脅迫下植物的化學防禦
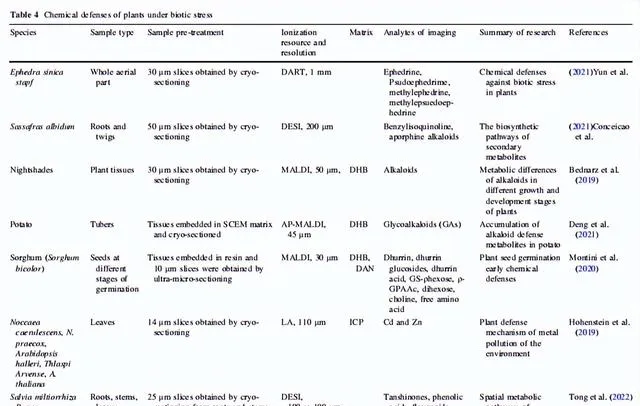
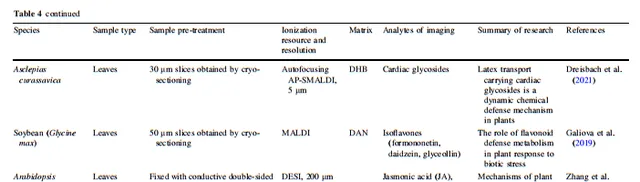
表5 植物根系與土壤的物質交換
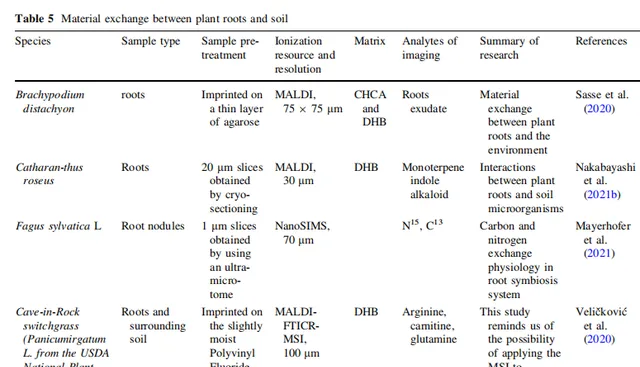
表6 植物-微生物相互作用
表7 植物次生代謝物的空間生物合成途徑
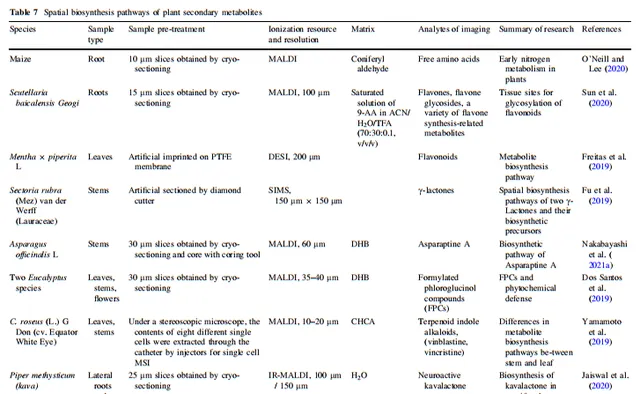

表8 外源物質在植物體內的代謝
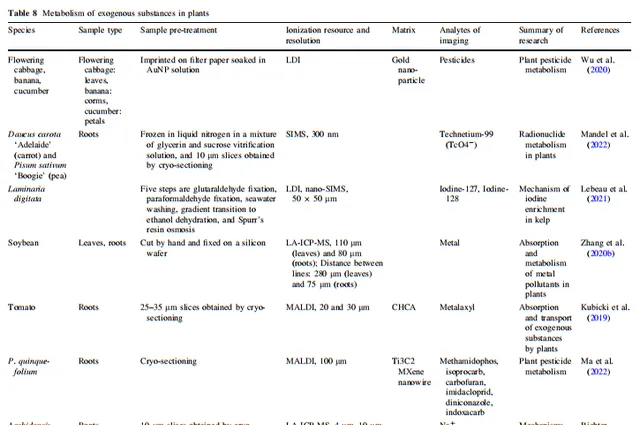
表9 其他套用
3.1 非生物脅迫下植物的脅迫耐受生理學
植物經歷了適者生存的自然選擇,呈現出一種區域性的生存趨勢。植物區域性生長的前進演化篩選考慮到一些非生物因素,如溫度、濕度、土壤鹽度、酸堿度以及金屬汙染。當這些非生物脅迫因素刺激時,植物微系統中發生的一些最重要的適應力反應會增強植物的脅迫耐受性,並被認為是植物代謝調控的一部份。植物中的這些多重化學防禦反應主要用於修復植物細胞的損傷,恢復植物細胞的穩態,並改變植物適應不同生長環境的方式。透過MSI在這些脅迫響應中重要的防禦代謝物的組織分布中,部份揭示了植物抵抗力的生理機制。
在幹旱、高鹽、強光或升高溫度的應對過程中,植物可以增加離層素的合成來促使葉片脫落並減少蒸散。然而,植物細胞的水勢由溶質勢、壓力勢和襯質組成,是理解植物抵抗幹旱機制的重要概念。面對各種非生物脅迫(如幹旱、過鹽和重金屬汙染土壤),植物透過增加小代謝物的生物合成,降低細胞的溶質勢並維持滲透壓,如在棉籽中觀察到的那樣。此外,在極端鹽堿或幹旱條件下,棉籽還透過增強甘油磷脂的生物合成來維持和補充細胞膜。在含有高汞和金含量的土壤中生長的木薯也出現了類似的情況;木薯在根表皮中具有相對較高的脂質含量。這些結果得到了Villette等人的研究的支持,該研究發現工業化合物及其代謝物主要被阻止在黑楊葉片靠近表皮的周圍組織中。此外,Gupta等人提出,植物細胞中黃酮類化合物的積累影響了植物滲透壓,並可作為耐鹽大麥品種中的代謝生物標誌物。
3.2 生物脅迫下植物的化學防禦
植物必須應對包括病原體侵蝕和動物咬食在內的生物壓力,以在生物圈中茁壯生長。在漫長的前進演化過程中,植物發展了多種次生代謝途徑,合成和積累一些獨特的防禦化學成分,用於組織中抵禦生物攻擊。植物學家利用MSI收集生物堿、黃酮類化合物等與防禦有關的化合物的空間代謝數據;然後利用這些數據解釋植物在生物壓力下的化學防禦。
堿類化合物在許多植物中是非常重要的防禦代謝物。這些堿類化合物儲存在特定的植物胞器中,受到脅迫訊號的刺激後被釋放並運送到作用位點,以抵抗真菌、細菌和其他病原體,以及昆蟲捕食者。已經證明,尼古丁、α-石竹烯和α-茄堿等堿類化合物對多種啃食性和吸食性昆蟲具有抵抗作用。士林和馬錢子堿是兩個堿類化合物的例子,它們與捕食者的酶結合,引起中毒或頭暈。根據Yun等人的MSI研究,麻黃的側枝中分布更多的麻黃堿和偽麻黃堿,這是因為前者更容易受到動物咬食。在檫木的地下部份,堿類化合物主要檢測到在根的外表皮中,用於抵禦草食動物、真菌、細菌等。然而,對於活體植物,特定防禦代謝物的積累可能在生長和發育的不同階段並不一致。Bednarz等人發現龍葵屬植物的主要堿類在不同的生長和發育階段發生了變化。隨著儲存時間的延長,馬鈴薯中四種有毒的配糖生物堿的合成增加,而在芽和果皮中的積累大於塊莖組織,這可能是馬鈴薯安全發芽的一種防禦機制。Montini等人發現dhurrin是一種重要的植物化學防禦化合物,其生物合成和內源代謝發生在種子早期發芽階段,以防止外界壓力。
對黃酮類化合物的空間代謝分析是研究植物化學防禦機制的另一個研究熱點。Sun等人發現,黃芩根部的外周韌皮部是黃酮類化合物合成和糖基化儲存的主要場所,在受到生物攻擊時能夠啟動快速的防禦反應,是其抵抗生物脅迫的重要生理表現。大豆和黃芩的防禦機制非常相似,都是在葉薄壁組織中積累類黃酮來抵抗蚜蟲。Tong等人發現 C-3 位點的羥基化啟用了脅迫下植物中類黃酮的重新分配。
除了積累防禦代謝物以防止損傷之外,植物還在受傷後經歷代謝調節變化,以抵抗捕食者或恢復穩態。Dreisbach等人揭示了當馬利筋葉片被動物咬食時,它透過分配和凝固橡膠乳液攜帶強心苷,以抵抗捕食者。植物激素也參與了植物受傷後的調節。Zhang等人發現茉莉酸(JA)、水楊酸(SA)、吲哚-3-乙酸(IAA)和離層素(ABA)都參與了植物葉片對機械應激的調節,其中JA的響應最高,而ABA沒有明顯的響應。SA的積累與JA及其前體化合物有關。
3.3 植物根系與土壤的物質交換
植物透過化學分子的語言與自然界進行交流,具體而言,植物主要透過與周圍空氣和土壤中的化學物質交換,與周圍環境不斷進行交流。土壤含有水分、許多元素和養分、豐富的微量元素,以及植物生長和發育所需的多樣有機物。植物與其環境之間的物質長期交換也是整個植物生命周期中植物分子生理學的主要組成部份。近年來,許多植物的物質交換過程已經透過MSI進行了確定。
植物的地下部份(主要是根系和可能的地下莖)是植物與土壤相互作用的主要場所。由於根尖缺乏卡氏帶和次生細胞壁,因此根尖是與周圍環境進行化學交換的主要場所;這是透過MSI的套用所推斷的,其空間分布結果顯示根分泌物在根尖最豐富,而在成熟和伸長區域則分布較少。此外,根分泌物在成熟和伸長區域的少量分布表明,也透過活躍的運輸或其他特殊通道發生了少量物質交換。此外,Veličković等人開發了一種根-土壤橫截面膜印法,以研究活躍的幼根與土壤之間的物質交換;然而,由於整體根體積的限制,他們的研究未發現常見的土壤有機化合物離子。他們檢索了整個根的空間代謝資訊,包括阿拉伯半乳糖、蘋果酸和其他關鍵代謝化合物,旨在確定植物對脅迫的抵抗基質。空間代謝組學在展示植物根部化學方面的潛力仍有待探索,主要是由於根-土壤橫截面成像的困難。
3.4 植物-微生物相互作用
最近,利用基於MSI的空間代謝組成像技術,深入探討了植物與微生物之間錯綜復雜的關系。不同的微生物群體以多種方式影響植物。Righetti等人使用AP-MALDI比較了福壽螺菌中各種代謝物的空間分布。他們還發現,亞油酸、α-亞麻酸及其相關代謝物在胚乳中積累,改變了其結構完整性。這一步是小麥福壽螺菌侵染的毒理學過程的一部份。然而,小麥中的許多防禦機制也被啟用,包括甘油三酯和半乳糖脂等訊號分子;這些分子參與植物細胞膜修飾、植物激素受體啟用、真菌辨識和植物創傷後反應。此外,細胞壁開始積累更多的羥基肉桂醯胺,透過多糖交聯與細胞壁成分結合,提高物理防禦屏障。
基於植物對致病菌的適應力,許多研究人員透過MSI探討了植物真菌的控制,為提高作物抗性提供了依據。例如,除了固氮作用之外,Hansen等人MALDI-MSI研究證實大豆結瘤還可以產生一些抗菌活性物質。Liu等人透過MALDI-MSI直接篩選了紫丁香中的抗菌活性物質,這為其他作物的真菌生物防治未提供證據。
MSI還為探索在一些植物中促進有益共生機制、促使微生物群落繁榮的機制提供了空間代謝資訊。Brescia等人使用模擬根際環境的生長培養基,證明植物透過釋放根際分泌物刺激Lysobacter capsici AZ78(AZ78)的細胞移動、生物膜形成和生物活性次生代謝產物的生物合成,從而影響土壤中的微生物群落,促進根系繁榮。對於促進生長的細菌,短柄草在根系中積累了類黃酮和酚酸等次生代謝產物,並增加了吲哚乙酸的合成以促進根系繁榮。由於MSI能夠提供精細的空間代謝資訊(植物與各種微生物以及微生物之間),因此在研究眾多植物與微生物相互作用方面具有重要的潛力。
3.5 植物次生代謝產物
植物的糖代謝、脂質代謝、蛋白質代謝、核酸代謝、生物氧化和氧化磷酸化等生理過程相互關聯,構成了維持植物正常生長和發育的基礎。植物在應對環境或生物脅迫時,演化出許多復雜的次生代謝途徑,形成了許多活性結構。植物的代謝網絡已經得到廣泛研究。許多類別天然產物的部份生物合成途徑已經被確定,並行展成完整的化學結構衍生途徑。MSI可以幫助將植物平面代謝網絡與植物形態特征相關聯,以理解植物的空間代謝特征。Yun等人檢測到麻黃側枝中麻黃堿和甲基麻黃堿等主要生物堿的分布高於主莖,而莖的內表面的分布高於外表面。這一結果可能與植物次生代謝產物對其自身的生物功能有關。O'Neill和Lee發現玉米根部的氮代謝在植物早期生長階段顯示出一定的親本遺傳性,透過獲取玉米根部遊離胺基酸的空間資訊,了解在環境影響之前植物氮代謝的模式。研究人員旨在簡化植物生長發育早期氮代謝(植物基本生理代謝)的研究,避免次生代謝的過度幹擾。Sun等人證明了根韌皮是黃酮合成和儲存的地方,也是黃酮苷酸的糖基化的地方。Li等人發現丹參根部的丹參酸在木質部合成,並部份儲存在栓質和栓皮中。然後,丹參酸透過維管束穿過莖,並部份保留在髓腔中,最終從葉脈擴散到葉肉。Freitas等人透過DESI-MSI觀察了一些在黃酮生物合成途徑中關鍵化合物,這些化合物有潛力將黃酮生物合成途徑與植物空間中的合成和代謝部位聯系起來。Fu等人推測五種γ-內酯可能是紅扇貝莖的射線薄壁細胞中同一生物合成途徑的一部份;他們的研究展示了TOF–SIMS MS/MS成像輔助其他技術了解植物內源產物生物合成途徑的潛力。Yamamoto等人透過代謝物的單細胞MSI發現了長春花的莖和葉之間的代謝產物的生物合成功能存在輕微差異。Revuru等人進一步提供了支持將skyrin作為金絲桃素前體化合物的證據,並透過MALDI-MSI假設後者的生物合成途徑發生在金絲桃葉的黑色腺體中,而大黃酚對金絲桃素的生物合成不直接相關,因為它在葉中廣泛分布。
3.6 植物體內外源物質的代謝
不同的植物可能對相似的外源物質擁有相似的代謝途徑。具體而言,植物在漫長的前進演化歷史中發展出代謝的多樣性,但仍保留了一些相似的基礎生理過程。例如,植物主要透過蒸散作用吸收中性化學物質和非必需金屬,因為植物吸收這些物量與蒸發的水量呈負相關關系。Kubicki等人利用MALDI-MSI證實了甲酯殺菌劑在番茄植物的常規吸水過程中是透過木質部和皮層完成系統傳輸的。同樣,農藥轉運體從韌皮到木質部擴散,隨著水在木質部中的運輸,最終在葉緣積累。這一動態過程由Wu等人透過MALDI-MSI進行了深入觀察。MSI分析還顯示,許多金屬汙染物仍停留在韌皮之外。此外,盡管鈉是植物的一種必需成分,但過多的鈉也被阻止進入韌皮部。然而,植物的代謝也存在一些變異性。Mandel等人的SIMS單細胞MSI數據顯示,植物對三種放射性核素的富集過程是不同的。鍀(Tc)透過主動運輸橫向轉移到木質部,並隨著水一起運輸,而錸(Re)被植物吸收較少,只在胡蘿蔔吸收後儲存並在薄壁組織中運輸。Lebeau等人透過DESI-MSI發現,在富含碘的海藻海帶中,放射性核素I和常見同位素I具有不同的細胞外分布。這些外源代謝數據的獲取為探索植物生理的外源調控提供了適當的基礎資訊。
3.7 其他套用
除了上述六大類套用之外,MSI還有一些其他的套用,這裏進行了總結。Witzel和Matros使用MALDI技術探索草莓代謝物與其外觀和口感之間的關系。De Moraes Pontes等人使用DESI-MSI尋找甜橙的病害標誌物,以實作對黃龍病的快速檢測。一些植物學家結合植物的空間代謝資訊與其生物學功能,解釋它們的代謝行為。綜上所述,MSI 在探索植物生理和病理機制方面顯示出顯著的套用潛力。
4 MSI的局限性和未來前景
回顧近期文獻,空間代謝組學能夠系統全面地展示植物代謝物的空間分布,有效地將植物代謝與植物形態聯系起來,從而為揭示植物分子生理學提供了豐富、多樣和高通量的數據支持。盡管透過MSI實作的代謝物的分子定位在植物空間代謝研究中並非不可替代,但其高通量的定位能力結合代謝組學和樣品處理的便利性在其他方法中是不可替代的。然而,不能忽視空間代謝組學的局限性。首先,並非所有植物組織都能夠進行空間分析,因為從一些脆弱、幹燥和堅硬組織中獲取適當樣本存在困難。其次,作為所有質譜技術的共同問題,離子抑制在MSI中也是一個重要考慮因素。因此,除了簡單的切片處理外,一些研究人員還使用有機溶劑的樣品預處理過程來去除雜質。然而,一旦預處理過程變得復雜,植物代謝物的變化就成為一個主要因素。第三,大量空間定位資訊的獲取是一個雙刃劍,給研究人員在提高植物研究系統性的同時提取植物生理資訊帶來了困難。最後,植物生理過程的變化只能根據空間代謝組學提供的實際植物代謝物來進行推理、假設或驗證。例如,許多植物代謝網絡操作的動態問題無法透過空間代謝組學解決。
我們認為,MSI的這些局限性不會掩蓋植物空間多組學研究模型在未來對植物生理特性、植物動態代謝、基因和可變表型之間的動態物質基礎進行研究的潛力。表型以及轉錄組、蛋白質組和代謝組變化的準確辨識和定位是植物獲得性遺傳研究的重要方面。透過多種高通量和高精度的組學方法,可以適當地表征植物的基因組、轉錄組、蛋白質組和代謝組。由於近距或遠距傳感工具的發展,現在可以精確分類植物表型。近距傳感是指測量靠近植物的器材,如電腦斷層掃描和磁共振成像,而遠距傳感則利用電磁波輻射、反射特性等方式遠離植物進行非接觸測量。MSI主導的空間多組學研究模型可以提供關於植物的跨組織和跨細胞代謝過程的資訊,促進植物結構、生理學、形態等復雜表型與轉錄組、蛋白質組和代謝組的多樣性之間關系的有效篩選。MSI主導的空間多組學研究模型預計將發展成為植物生理學解釋和育種設計最佳化的有力工具,以及探索食用和藥用植物農學和植物生理學的有力工具。











