文章1

低氧應答途徑調控蘭科植物鐵皮石斛原球莖與真菌的共生
期刊:
IF: 7.4
發表時間: 2023年3月
單位: 雲南大學
研究背景
雲南大學生態與環境學院生物多樣性研究院韓佳嘉和高江雲團隊在線上發表了題為「Symbiosis between protocorms and involves the plant hypoxia response pathway」的研究論文,該研究利用轉錄組,基因功能驗證等方法揭示了真菌是一種能與多種蘭科植物建立菌根伴侶關係的內生真菌,並行現植物ADH(乙醇去氫酶)及其相關的低氧應答途徑調控了蘭科植物鐵皮石斛原球莖與真菌的共生。 邁維代謝為其提供了轉錄組檢測與分析服務。
菌根是真菌和植物根之間普遍存在的共生體。蘭科植物種子細小,胚未完全分化、幾乎沒有胚乳,需要依靠特定的共生真菌提供營養來促進其萌發和後續原球莖的生長。不同於有廣泛宿主範圍的叢枝菌根真菌,蘭科植物真菌對他們的宿主通常具有高度特異性。然而,與叢枝菌根和根瘤菌共生的分子機制相比,蘭科菌根共生的分子機制尚不清楚。在本研究中,作者發現內生真菌是一種廣譜的蘭科植物親和性真菌,能夠促進多種(2個屬6個種)附生蘭科植物的種子萌發並促進其原球莖發育成幼苗,其中包括鐵皮石斛()。鐵皮石斛原球莖與真菌的共生,為後續研究提供了重要模型。
研究材料
12種生態和分類學上不同的蘭科種子,無菌和共生鐵皮石斛原球莖。
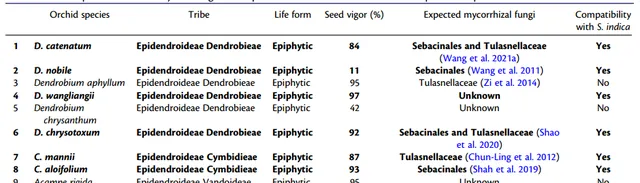
技術路線
研究結果
1、能夠與鐵皮石斛()及其他蘭科植物共生
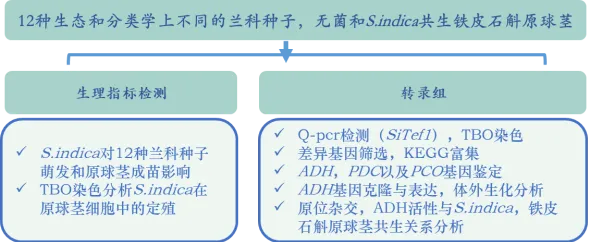
作者分析了內生真菌對收集的12種生態和分類學上不同的蘭科種子萌發和原球莖成苗的影響。結果發現,促進了其中6種的生長:鐵皮石斛()、金釵石斛()、王亮石斛()、鼓槌石斛()、紋瓣蘭()和硬葉蘭()(圖1A,B)。不僅刺激了這些種子的萌發,而且促進了原球莖的發育(圖1A,B)。對鐵皮石斛的促生長作用較強,但對硬葉蘭較弱(圖1B)。作者透過半薄組織切片甲苯胺藍(TBO)染色分析了在原球莖細胞中的定殖。細胞內菌絲體位於蘭科植物原球莖的基部區域(圖1C),該區域為未來根系發育的位置。細胞內菌絲團(圖1C)在原球莖中的形態與其他蘭科菌根(OrM)一致。因此,該研究結果表明,與一些蘭科植物形成了典型的OrM,並促進了其種子萌發和原球莖的發育。在這些蘭科植物中,鐵皮石斛是一種具有較高藥用價值的瀕危植物,並且已經公布了參考基因組。因此作者利用鐵皮石斛-共生系統研究了蘭科植物與內生真菌之間的共生機制。
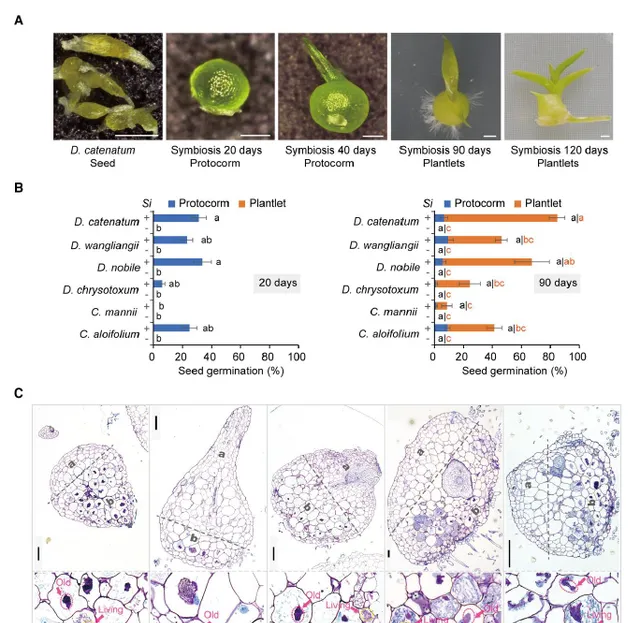
圖1 與多種蘭科植物共生。A:鐵皮石斛種子和原球莖在接種後的形態;B:(Si)促進6中蘭科植物種子萌發(接種約20d後形成原球莖)和原球莖發育(大多數原球莖在90天內發育成小植株);C:半薄組織切片甲苯胺藍(TBO)染色的原球莖切片,(a)蘭科原球莖的頂端未感染區和(b)基部菌根區,底部為定植細胞的細節。
2、共生原球莖與無菌原球莖的表達譜差異分析
在這項研究中,為了獲得足夠的樣本量用於RT-qPCR和轉錄組分析,作者使用了無菌萌發種子30天齡的鐵皮石斛原球莖作為原材料,將30天大的原球莖移植到共生培養基(OMA培養基)中進行無菌培養(CK)和接種(Si),然後收集用於隨後的RT-qPCR和轉錄組分析。對在接種後的原球莖中的轉譯延伸因子1()表達進行RT-qPCR分析,以鐵皮石斛Actin-7()為內參,RT-qPCR結果顯示定殖水平在接種後第9天達到峰值(圖2A)。半薄組織切片甲苯胺藍(TBO)染色結果也證實了共生原球莖中存在細胞內菌絲體,位於原球莖的基部區域(圖2B),這與前期觀察結果一致(圖1C)。因此在接種後第9天的原球莖和真菌之間建立了穩定的伴侶關係。作者選擇接種後第9天的共生原球莖用於隨後的轉錄組分析。
轉錄組分析顯示,無菌(control)和共生原球莖()的表達譜不同(圖2C,D)。作者從vs control中篩選鑒定出356個差異表達基因(DEG)(log2(fold change)≥1,q≤0.05),KEGG通路富集發現,這些DEGs主要富集在植物防禦和免疫應答相關通路(圖2E),包括植物激素訊息傳遞、植物病原體互作和MAPK訊號通路,除此之外還包括類黃酮生物合成、苯丙素生物合成和色胺酸代謝通路。在大多數富集途徑中,上調和下調的DEG數量相似(圖2E)。觀察到第18天的定殖水平低於第9天(圖2A),作者認為在穩定的菌根伴侶關係建立後,菌根真菌在寄主植物中的定殖是有限的,並且這種定殖影響了蘭科植物的防禦和免疫相關途徑。此外,糖解作用途徑4個基因在糖解作用途徑中上調,推測ADH在鐵皮石斛菌根共生中起著重要作用。
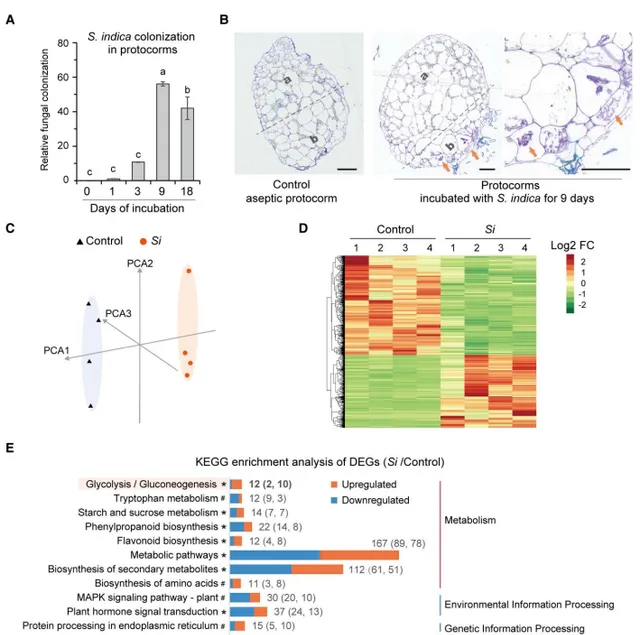
圖2 共生原球莖轉錄組分析。(A)的RT-qPCR分析;(B)無菌和共生原球莖頂端區域(a)和基底區域(b)的半薄組織切片;(C)轉錄組數據PCA分析;(D)聚類熱圖分析;(E)差異表達基因(DEGs)的KEGG富集分析。
3、低氧適應力基因在共生原球莖中被誘導
ADH和PDC(丙酮酸去氫酶)在植物低氧反應過程中發揮著重要作用。透過同源性分析,作者鑒定出12個同源基因。轉錄組和RT-qPCR分析顯示,在共生原球莖中,幾乎所有的基因都被上調,其中和上調最顯著(圖3A,C)。生化分析表明接種後第9天,共生原球莖中ADH的還原活性明顯高於無菌原球莖,與上述結果一致(圖3D,E)。這些結果表明,和可能與菌根共生特異性相關。此外,基因在共生原球莖中顯著上調(圖3B,C)。缺氧誘導基因和也上調(圖3B,C)。此外,和等低氧適應力基因的表達與共生時間呈正相關。這些結果表明,的定植能夠誘導鐵皮石斛共生原球莖中低氧適應力基因(特別)的表達。
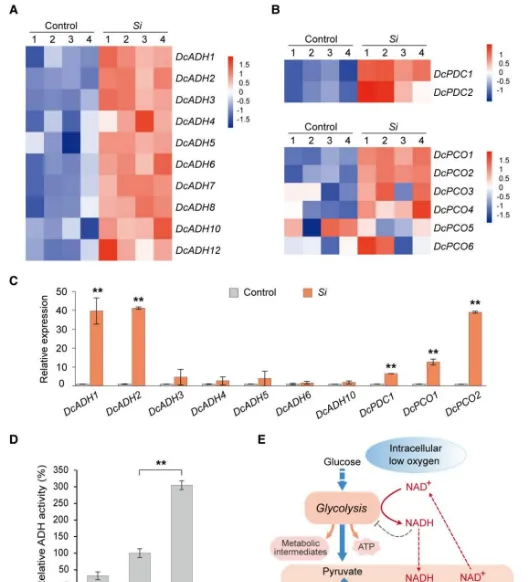
圖3低氧適應力基因在共生原球莖中被誘導。(A、B)和基因表現熱圖;(C)關鍵的低氧適應力基因的RT-qPCR分析;(D)無菌原球莖以及共生原球莖中ADH活性;(E)植物缺氧代謝受ADH和PDC調控。
4、ADH活性正向調節原球莖菌根共生
為了驗證的上調是否與共生原球莖中ADH活性升高有關,作者複制了和的編碼序列,並在大腸桿菌中表達。重組菌蛋白萃取物的體外生化分析顯示,和具有預期的ADH活性,可將NADH轉化為NAD+(圖3E和4A)。接著作者證實了葛根素和4-甲基吡唑(ADH抑制劑 )均抑制了和的NADH轉化活性(圖4B,C)。其中在共生原球莖中表達量最高。因此作者使用地高辛(DIG)標記的寡核苷酸探針進行了原位雜交。在共生原球莖中,主要在菌根基部表達,而不是在根尖表達(圖5A,B),這與在原球莖中的定殖位置一致。接著作者使用葛根素來抑制的功能,並觀察其對與鐵皮石斛原球莖伴侶關係的影響。作者發現葛根素顯著降低了在鐵皮石斛原球莖上的定植(圖5C),但對的生長沒有顯著的不利影響(圖5D)。另一種ADH抑制劑4-甲基吡唑也顯著降低了在鐵皮石斛原球莖上的定植,但在200 μM濃度下,4-甲基吡唑能輕微抑制的生長。這些結果表明,抑制ADH活性可以抑制在鐵皮石斛原球莖上的定植。綜上結果表明,ADH及其相關的低氧應答途徑調控蘭科植物鐵皮石斛原球莖與真菌的共生。
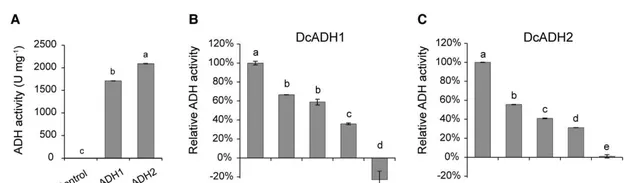
圖4 和蛋白的ADH活性。(A)體外實驗表明,純化後的和蛋白具有ADH還原活性,可催化NADH轉化為NAD+;(B, C)不同濃度葛根素對純化後的和還原活性的抑制作用。

圖5 ADH活性是共生原球莖的OrM真菌定殖所必需的。(A) mRNA在無菌和共生原球莖的縱向切片上的原位雜交;(B)正義(而非反義)探針的原位雜交;(C)體內試驗表明,在接種後第9天,葛根素抑制了在共生原球莖中的定殖;(D)葛根素對的生長沒有產生顯著的不利影響;(E)ADH調節蘭科原球莖中菌根共生的模式圖。
文章2

轉錄組+代謝組解析人參皂苷脅迫誘導三七根分泌物變化以招募有益的伯克霍氏菌株B36
期刊: J. Agric. Food Chem.
IF: 6.1
發表時間: 2023年3月
單位: 雲南農業大學
研究背景
雲南農業大學植保學院楊敏老師團隊在J. Agric. Food Chem.線上發表了題為「Autotoxic Ginsenoside Stress Induces Changes in Root Exudates to Recruit the Beneficial Strain B36 as Revealed by Transcriptomic and Metabolomic Approaches」的研究論文,該研究利用轉錄組和代謝組等方法闡明了人參皂苷脅迫啟用了苯丙素生物合成和α-亞麻酸代謝通路,從而增加了肉桂酸、2-十二烯酸和12-氧代植物二烯酸(12-OPDA)的分泌。其中肉桂酸可以同時促進B36的趨化和生長,增強B36在根際定殖,最終提高三七的成活率。這一發現將促進有益菌在農業生產中的實際套用,並透過外源添加關鍵代謝產物實作可復制的生物防治效果。 邁維代謝為其提供了代謝組和轉錄組檢測分析服務。
三七((Burk.)F.H.Chen)由於土傳病原菌和自毒性物質人參皂苷的積累,易患嚴重的根腐病。前期研究表明,在人參皂苷自毒脅迫下,三七能在根際土壤中富集有益的伯克霍爾德菌B36,該菌能降解自毒皂苷 同時拮抗土傳病原菌 ,最終緩解根腐病,這為透過控制根際微生物來克服根腐病提供了一種新的生態策略。然而,自毒性物質人參皂苷是否能誘導三七根系轉錄組和分泌物的變化,進而募集有益微生物,目前尚不清楚。
研究材料
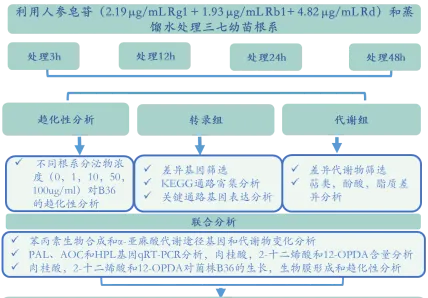
三七幼苗在人參皂苷(2.19 μg/mL Rg + 1.93 μg/mL Rb+ 4.82 μg/mL Rd)溶液和蒸餾水溶液中培養3、12、24和48h,在每個時間點下收集根組織進行轉錄組分析並收集玻璃瓶中的溶液,用濾紙和0.22 μm親水性膜過濾後濃縮成幹物質進行代謝組分析。每個處理3個生物學重復。
技術路線
研究結果
1、人參皂苷脅迫下三七根系分泌物可吸引伯克霍爾德菌B36
當三七根受到人參皂苷脅迫後,三七根系分泌物對B36表現出明顯的吸引作用,且呈劑量依賴性(圖1))。具體而言,當分別利用脅迫處理3h,24 h下濃度為10、50和100 μg/mL的三七根系分泌物濃度以及12 h下濃度為100 μg/mL處理時對菌株B36表現出顯著的吸引作用(圖1A,B,C)。人參皂苷脅迫48 h後,與無菌水對照相比,根分泌濃度為50 μg/ mL時對菌株B36具有顯著的吸引作用(圖1D)。此外,人參皂苷滅菌水處理與對照之間無顯著差異。上述結果表明,在人參皂苷脅迫下,三七根系可以分泌一些代謝產物來吸引B36。
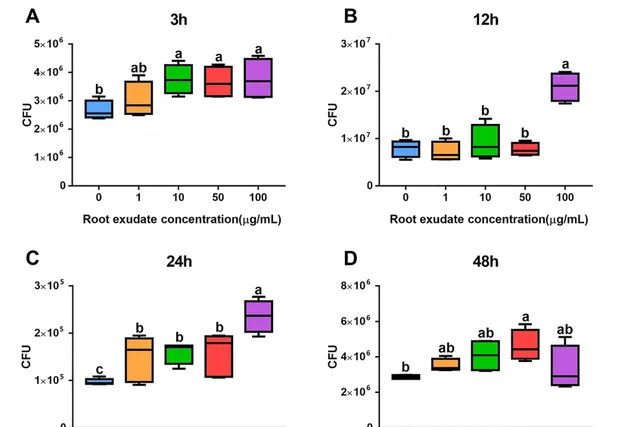
圖1 人參皂苷脅迫處理3 (A)、12 (B)、24 (C)、48 h (D)時不同濃度根分泌物對伯克霍爾德菌B36的趨化性測定。
2、人參皂苷脅迫處理改變三七根的轉錄組譜
利用人參皂苷和蒸餾水分別脅迫處理3、12、24和48 h的三七根進行轉錄組分析。共24個文庫(ck-3h, g-3h, ck-12h, g-12h, ck-24h, g-24h, ck-48h, g-48h,每個處理3個生物重復)得到約159.62 Gb的clean bases,Q20和Q30的平均值分別為98.11和94.32%,平均GC含量為43.66%。利用NR、GO、KEGG、Pfam、KOG和SwissProt數據庫註釋後進行差異基因篩選。4個比較(ck-3h .g-3h, ck-12h .g-12h, ck-24h .g-24h, ck-48h .g-48h)分別篩選到33、15、713和126個DEG。其中分別有25、5、615和106個DEG在人參皂苷處理時顯著上調(圖2A)。說明人參皂苷脅迫24 h後,根內基因表現水平發生了顯著變化。KEGG通路富集分析結果顯示,在人參皂苷脅迫3、12、24和48 h後,分別有1、1、9和2個通路被顯著富集(圖2B),其中苯丙素生物合成途徑在脅迫3和12 h下均顯著富集(圖1)。在脅迫24 h後顯著富集的9條途徑中(圖2B),苯丙素生物合成和二芳基庚烷和姜醇生物合成途徑的DEGs大部份上調表達(圖2C)。在苯丙素生物合成過程中,大部份編碼過氧化物酶(POD)、反式肉桂酸4-單加氧酶(CYP73A)、β-葡萄糖苷酶、反式肉桂酸-4-羥化酶(CYP98A)、莽草酸羥基肉桂酰基轉移酶(HCT)、肉桂醇去氫酶(CAD)和東莨菪素葡萄糖轉移酶(TOGT1)的基因在人參皂苷處理24 h後顯著上調,其中編碼CYP73A和CYP98A的基因(Pno05G006544和Pno01G007403)在人參皂苷處理24 h後分別上調了2.99倍和3.43倍。二芳基庚烷和姜醇生物合成中編碼反式白藜蘆醇二-O-甲基轉移酶(ROMT)的基因也顯著上調。另外在富集到α-亞麻酸代謝、花生四烯酸代謝及其上遊途徑不飽和脂肪酸生物合成和脂肪酸代謝中的DEGs均上調(圖2C)。例如編碼FAD2、ACSL、TGL4、CYP4A、DAD1、OPR、DOX、HPL的基因。其中,編碼OPR的基因(Pno02G004829和Pno02G004840)在人參皂苷處理24 h後分別上調了3.48倍和3.47倍。這些結果表明,參與上述途徑的DEG可能與三七根對人參皂苷脅迫的反應密切相關。
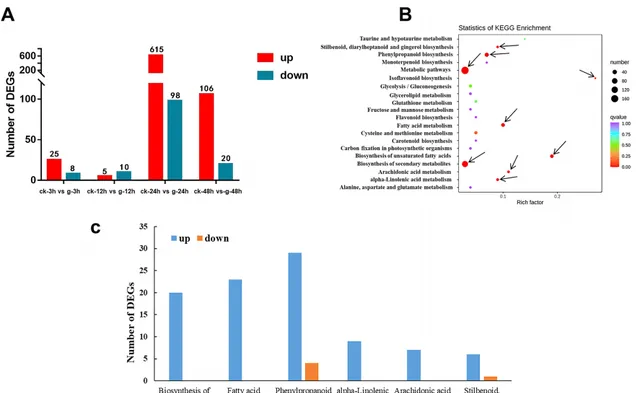
圖2 人參皂苷脅迫對三七根轉錄組影響。(A)上調和下調基因柱狀圖統計;(B)人參皂苷處理24 h後KEGG富集分析;(C)人參皂苷處理24 h後顯著富集途徑中的DEGs數量。
3、人參皂苷脅迫下根系分泌物代謝組學分析
在所有樣品中總共檢測到273種代謝物,PCA結果顯示脅迫不同時期根系分泌物代謝譜與對照組差異顯著,且不同時期間也明顯分離。透過VIP≥1和|log2(FC)|≥1篩選差異代謝物(DAM),共得到250個DAM,其中人參皂苷脅迫處理3、12、24、48 h時,代謝物分別上調34、31、28、22個,下調52、27、41、15個。根分泌物中響應人參皂苷脅迫的DAM主要分為7類,包括胺基酸及其衍生物、酚酸、核苷酸及其衍生物、萜類、有機酸、脂類等(圖3A)。在人參皂苷處理3、12和24h後,萜類化合物(人參皂苷Rg1、人參皂苷Rp、三七皂苷A4、三七皂苷R1和三七皂苷R2)均顯著上調(圖3A)。此外,人參皂苷處理24h後,酚酸(肉桂酸、4-甲氧基桂皮醛、2胺基-3-甲氧基苯甲酸等)和脂質(2-十二烯酸、12-OPDA等)顯著上調,而有機酸(2-氧庚二酸、2-甲基-3-氧己二酸等)顯著下調(圖3B)。因此作者推測人參皂苷脅迫可能導致三七根分泌物中萜類、酚酸和脂類積累增加。
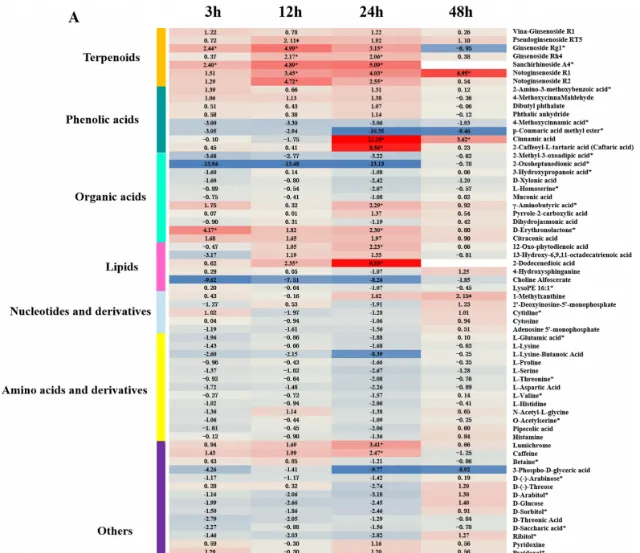
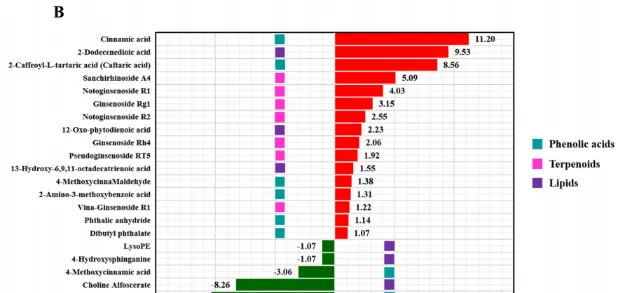
圖3 人參皂苷脅迫下代謝物差異分析。(A)人參皂苷處理不同時期代謝物顯著變化熱圖;(B)人參皂苷處理24 h後酚酸、萜類和脂類變化條形圖。
透過結合轉錄組和代謝組聯合分析(圖4A),作者選擇苯丙素生物合成和α-亞麻酸代謝途徑中顯著變化的基因,包括編碼PAL、AOC和HPL的基因進行qRT-PCR分析(圖4B)。同時作者檢測了人參皂苷處理24h後肉桂酸,2-十二烯酸和12-OPDA含量變化,發現處理後三七根分泌物中肉桂酸含量顯著增加至2511.194 ng/g,2-十二烯酸和12-OPDA含量顯著降低(圖4C)。
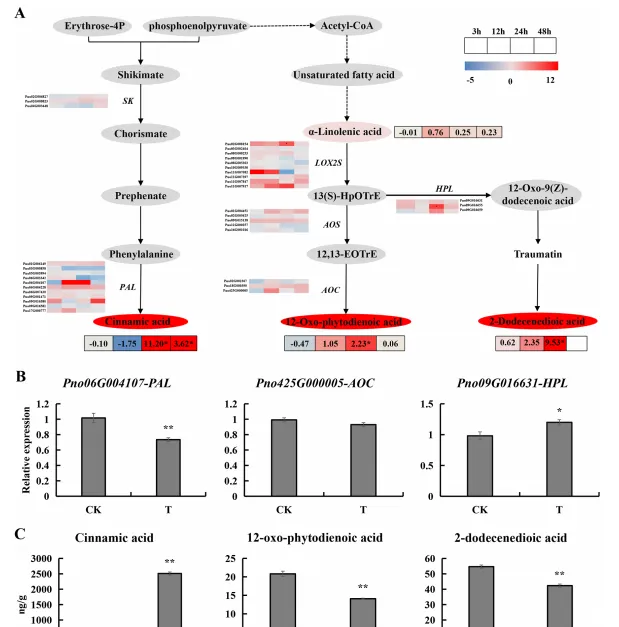
圖4 人參皂苷脅迫3、12、24和48 h後苯丙素生物合成和α-亞麻酸代謝相關基因和代謝物表達譜;(A)苯丙素生物合成和α-亞麻酸代謝通路熱圖;(B)PAL、AOC和HPL基因表現量;(C)肉桂酸、2-十二烯二酸和12-OPDA含量。
4 人參皂苷刺激三七根系分泌物中代謝物積累以增強B36定植
研究發現,人參皂苷脅迫促進三七根中肉桂酸分泌增加。趨化性實驗數據顯示,低濃度(1、10和50 μg/mL)的肉桂酸對菌株B36有明顯的吸引作用(圖5A),且在10和100 μg/mL濃度下顯著促進了菌株B36生物膜的形成(圖6A),在50μg/mL濃度下培養36 h時顯著促進了菌株B36細胞的生長(圖6D)。然而,高濃度的2-十二烯酸和12-OPDA(100 μg/mL)對菌株B36具有驅避性(圖5B,C)。2-十二烯酸對生物膜的形成沒有明顯的促進作用(圖6B)。這些結果表明,肉桂酸可能是根際B36趨化的重要訊號物質。
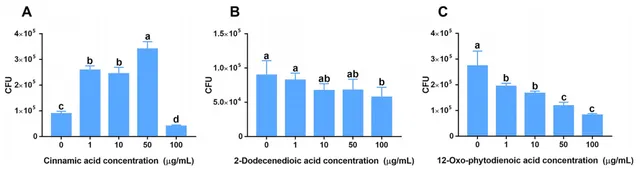
圖5伯克霍爾德菌B36對肉桂酸(A)、2-十二烯二酸(B)和12-OPDA (C)的趨化反應。
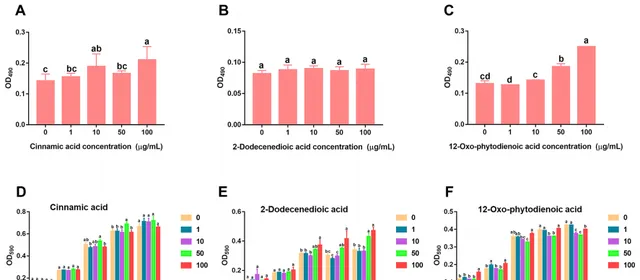
圖6 肉桂酸、2-十二烯酸和12-氧-植物二烯2-十二烯二酸(B)和12-OPDA對伯克霍爾德菌B36生物膜形成(A−C)和生長(D−F)的影響。
結合以上結果,作者進一步檢測了肉桂酸對B36在無菌土壤中定殖能力的影響。利用三七根分泌物和肉桂酸處理B36土壤,在處理3、5、10 d後,B36的定殖能力顯著增強。更重要的是,與根系分泌物處理相比,肉桂酸處理下B36在無菌土壤中的定殖數量顯著增加(圖7A)。盆栽試驗進一步證實,在連作土壤中單獨添加肉桂酸和B36後,對三七的存活率並沒有明顯的促進作用,但肉桂酸與B36配合使用,可以顯著提高三七在連作土壤中的存活率(圖7B)。
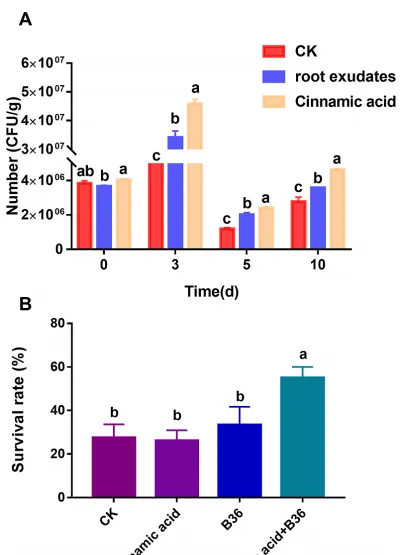
圖7 三七根分泌物和肉桂酸對無菌土壤中B36定植的影響(A),肉桂酸和B36對連作土壤中三七成活率的協同效應(B)。
往期熱門分享














