前言
花生瘡痂病是中國花生產區重要的真菌病害,產生光敏性真菌毒素痂囊腔菌,具有強烈的細胞毒性,為重要的毒力因子,前期研究發現,ESC生物合成極顯著地受到光照調控,揭示病菌毒素生物合成的光調控作用及其分子機制,對探究該病菌的侵染過程、致病機制和調控網絡具有重要意義,可為病害防治策略的高效制定提供了科學理論支撐。
本文開展了光質對病菌生長發育、ESC毒素產生及相關基因表現的影響效應研究,透過光質培養下樣品進行了轉錄組測序及光受體基因挖掘,對花生瘡痂病菌藍光受體EaWC1基因複制及生物資訊學分析,構建了基因剔除載體並初步驗證了藍光受體EaWC!基因功能,研究結果旨在為花生瘡痂病菌致病機制解析,探究光調控網絡提供重要分子證據,為病害綜合防治策略制定奠定科學的理論基礎。

光質對真菌的調控作用
近年來研究表明,光質對真菌生長發育及次生代謝產物的影響極其顯著,作為模式真菌粗糙脈孢菌的藍光響應包括誘導孢子形成和性發育和菌絲體類胡蘿蔔素的合成。
與黑暗相比,白光和藍光可以抑制輪枝鐮孢菌)、早疫病菌和玉米灰病菌的分生孢子形成,促進生長和毒素含量產生,而黑暗和紅光抑制構巢曲黴和竹黃菌分生孢子的產生,促進曲黴毒素和竹紅菌素的產生。
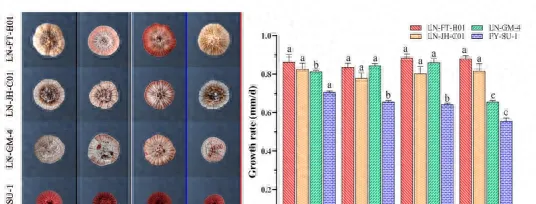
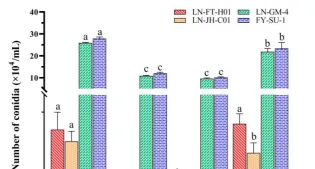
黑暗時灰黴病菌對葡萄和番茄的致病力最強,白光及藍光是致病力較弱,反而綠光對其在葡萄和草莓上致病力具有很強的抑制作用,不同光質下培養5株不同的性狀的蛹蟲草菌株,均表現為藍光抑制其菌落生長,白光和藍光極顯著促進分生孢子的產生。
黑暗條件下僅產生少量分生孢子,真菌光質響應的研究揭示了一種意想不到的復雜性,不同物種間對光質響應存在很大差異。
藍光受體
真菌對450nm(藍色)至700nm範圍內波長的光有光響應,藍光受體更易受光刺激產生光訊號,快速啟動光感系統,調控下遊次生代謝產物編碼基因表現而備受關註,目前已經在真菌中鑒定了幾類藍光受體,包括WCI,WC2,VIVID,隱花色素以及光解酶。
粗糙脈孢菌中真菌光受體的分子機制已在得到了較為詳細的研究,最早從粗糙脈孢菌中複制得到第一個真菌藍光受體基因WCI,該受體對N.crassa晝夜節律、生長趨向性、無性發育、有性生殖、抗UV損傷和類胡蘿蔔素等具有調控作用。
目前在構巢曲黴、煙曲黴、禾谷鐮刀菌、卷枝毛黴菌、深綠木黴、交鏈格孢、灰黴菌和玉米灰斑病菌等多種真菌中均篩選鑒定到藍光受體ⅣC1同源基因。
真菌光受體在結構上擁有多功能結構域,藍光受體WC1編碼蛋白通常具有三個PAS結構域和一個鋅指結構域,其中兩個PAS域用於蛋白質-蛋白質互作,鋅指結構域用於核定位,靠近N端的PAS域屬於一類特殊的結構域,稱為LOV結構域,也稱光、氧或電壓域。
之後不久克降了藍光檢測所必需的第二種蛋白質,命名為WC2,具有一個鋅指結構域和單個PAS結構域,可定位於細胞核,WC蛋白對於真菌光響應具有調節周期節律,促進或抑制有性發育,分生孢子產生及次生代謝產物合成積累的功能。
VIVID是一種小蛋白質簡稱VVD,具有一個與FAD或FMN結合形成的LOV結構域,在光照後基因表現的瞬間啟用中起重要作用,與WC1和WC2不同的是,在脈孢菌核中沒有發現VVD,只有在光誘導後才能在細胞質中觀察到活性物質,VIVID對生物鐘啟動極為重要,光訊號誘導生物鐘啟動會調節生物內部從而適應生長環境。
接收藍光到藍紫波段光訊號的受體還有隱花色素,它在原核和真核生物中均被發現,其廣泛的存在於構巢曲黴、粗糙脈孢菌、玉米灰斑病菌、木黴和核盤菌等多種真菌中,其重要調控真菌的光修復,生物鐘,有性無性發育和次生代謝等。
光敏色素最先是在植物中發現的,人們對其進行了充分的研究,在真菌中,這種光感受需類別首先在兩種模型真菌構巢曲黴和粗糙脈孢菌中被發現,目前關於真菌紅光受體的研究還主要集中於曲黴中,其他真菌中也被發現如灰黴崗。
光敏色素是一種大型的蛋白質,由一個結合發色團的光感區和一個訊號輸出域組成,感覺區域由三個區域組成域,GAF小配體結合區域和PHY光敏色素域。
面訊號輸出區域與原核組胺酸激相似,包括HKD組胺酸激酶結構域,HATPase腺苷三磷酸酶結構域,RRD響應調控域,光敏色素編碼基因Fph同樣調控真菌重要的生理生化過程,目前的研究主要集中在有性無性發育和色素沈著上。

盡管已經證明構巢曲黴中FphA可以使用膽綠素,但真菌光色素中膽紅素發色團衍生物的性質仍有待確定,發色團與PAS或GAF域結合並存在於兩種可互換的構象中,它們的光譜特性很好地區分開案,光敏色素以P形式吸收紅光,從面改變構象並將吸收最大值移至遠紅光光譜,這個新形式叫做p,在吸收遠紅光時,發色團恢復到Pr形式完成光響應迴圈。
藍光受體對真菌次生代謝的調控機制
細胞生物中光感應與傳導是一個復雜的生物過程,作為對真菌影響最廣泛顯著的藍光,藍光受體是各種真菌生長發育和次生代謝等生理生化過程所必須的生物感受器,眾多學者對藍光受體功能進行研究。
光生物學研究表明,藍光受體WCI基因的表達調控與老種真崗次生代謝產物的生物合成具有高度關聯性,光照能夠促進水積惡苗病菌中鏡孢菌素的生物合成,此時藍光受體co4基因高表達,模式真菌粗糙脈孢菌中C-1基因影響生物鐘和類胡蘿蔔素的生物合成。
禾谷鑲抱菌中藍光受體Fgwcl基因卻主要影響有性繁殖和無性發育,灰黴菌中紅光受體光敏色素Bqphy2,Bcphy3基因的缺失均抑制黑色素的生物合成,但突變株的致病性卻不相同,在缺失2基因的突變體中致病力顯著增強,面缺失Bcpp基因的突變株致病力明顯降低。
鏈格抱菌中藍光受體De基因強烈誘導了毒素的生物合成,但負調節Alternariol(AOH)毒素產生,玉米尾孢菌中藍光受體基因CRPI表現出促進毒力因子尾孢菌素積累,CRPI基因缺失明顯降低病菌致病性和病斑擴充套件能力。
雪洲鏡孢菌的研究中發現,藍光受體FamC!在感知光訊號、調節抗紫外線(UV)損傷、次生代謝、有性生殖及致病力中同樣是必需的,該基因的缺失導致光誘導的光解酶基因表現的缺失,從面導致對紫外線的過敏。
藍光受體用C1在真菌生長發育、次生代謝和環境感知中所行使功能具有復雜性和多樣性,調控模式和機理亦相差甚遠,1.3.2細胞中其它光訊號轉換。
菌株培養於PDA平板上,置於25℃光照培養,培養時間分別為14d、21d、28d.35d和42d,隨後打取菌餅,烘幹後以丙酮為提取溶劑萃取,采用超聲振蕩法提取至無色後合並,減壓濃縮後為病原菌毒素粗提液。
定量檢測參考劉璐(劉璐等,2018)方法,以紫外分光光度法繪制標準曲線,采用標準曲線法定量毒素,選擇最適培養天數,進行後續毒素測定的試驗。
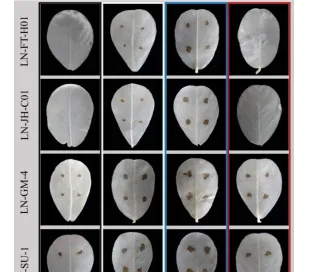
采用菌懸浮液定點定量離體接種法接種菌株,根據2.1.2制備菌懸液,取苗後14-28d健康無病,長勢均勻,展開的花生葉片,用無菌水輕拭葉片,對花生離體葉片進行保濕培養。
在葉片設定4-6個對稱的接種點,以無菌水為對照,每接種點接種20L菌懸液,置於不同光質條件下,22-25℃保濕培養,每日觀察發病情況,接種15d時拍照,脫色記錄病情。
差異表達基因分析
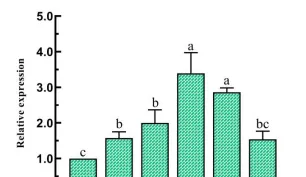
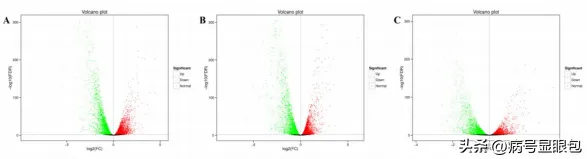
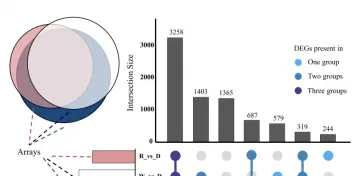
以同條件下黑暗處理的花生瘡痂病菌作對照,分析不同光質條件處理36h後的差異表達基因,WVSD共有5926個差異基因,其中2877個基因上調,3049個基因下調:BvsD共篩選出6344個差異基因,其中3096個基因上調表達,3248個基因下調表達:RvSD共有4507個差異表達基因,其中2094個基因上調,2413個基因下調。
說明本研究中不同光質處理下花生瘡痂病菌的基因表現受到調控,處理中差異表達基因數量關系,與黑暗相比,只在藍光處理時,顯示的差異表達基因有1365個,只在白光處理時,顯示的差異表達基因有579個,只在紅光處理時顯示的差異表達基因有244個。
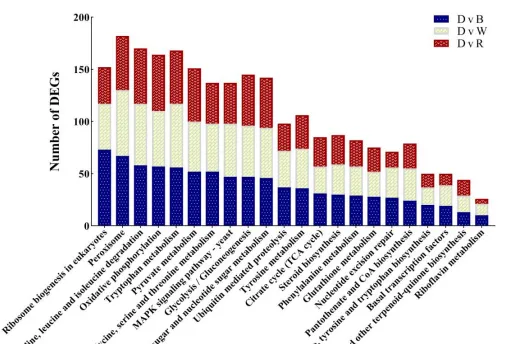
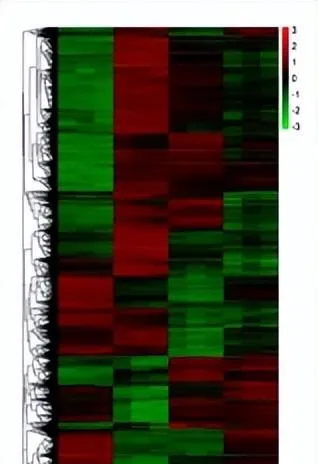
白光和藍光處理下差異表達基因遠遠大於紅光處理時數量,表明與黑暗相比,白光和藍光處理誘導基因的表達比紅光處理時多,藍光條件處理下誘導的基因表現最多,差異表達基因聚類分析圖中也存在相似的趨勢,差異表達基因上調倍數越大,顏色越紅,反之越綠。
黑暗處理時上調的基因,在白光和藍光處理時下調黑暗處理下調的基因,在白光和藍光處理時上調,紅光(R)處理時基因表現與黑暗基因一致,這表明白光和藍光處理與黑暗對照處理之間存在很大的差異,在紅光處理時的基因表現和黑暗處理相似。
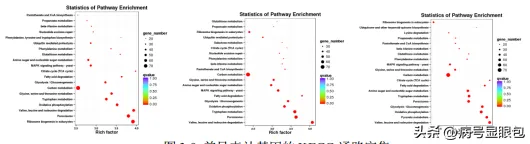
結論
為探究光對於花生瘡痂病菌基因表現的調控,共篩選到474個受光質誘導表達的基因,包括11個假定光受體基因,57個轉錄因子,81個參與光訊號通路基因,352個與次生代謝相關基因,透過Blast比對及系統發育樹分析表明,EVM0003940為WC-1同源基因,EVM0001480,EVM0004523,EVM0002126和EVM0003727均為隱花色素同源基因。
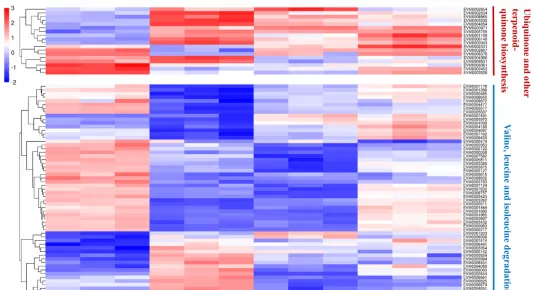
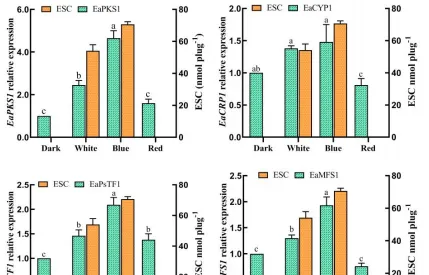
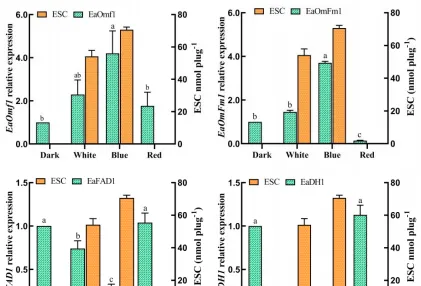
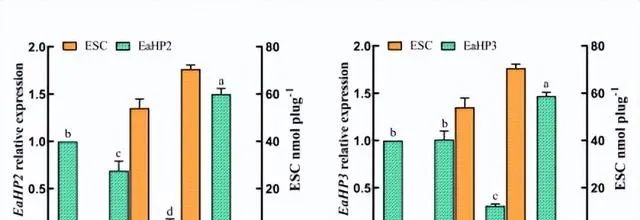
不同光質條件對花生瘡痂病菌ESC毒素產生具有顯著影響,對代謝途徑中轉錄組數據的DEGs分析有助於辨識花生瘡痂病菌花醌類化合物通路中基因在不同光質處理下的表達模式,透過Blast對不同光質處理下的DEGs進行功能分析。
預測325個可能是參與次生代謝產物生物合成的基因,根據這些基因的功能註釋,主要集中在9個功能組,分別為聚酮合酶、非核糖體肽合成酶、FAD依賴性單加氣酶、甲基轉移酶、FAD依賴性氣化還原酶、NADPH去氫酶、轉運蛋白、細胞色素P450和鐵還原酶類蛋白。
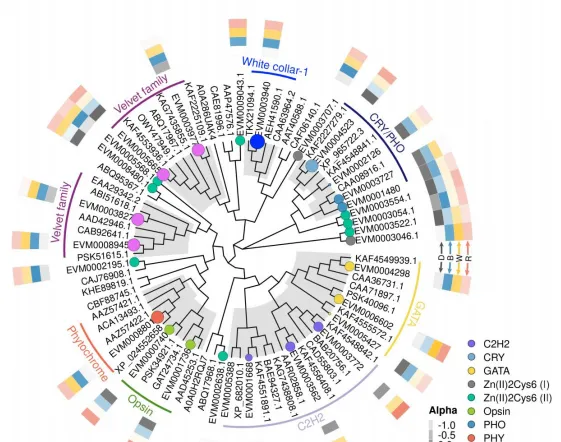
透過Blast和系統發育分析,成功篩選出12個次生代謝相關基因,透過熒光定量PCR表達驗證,結果表明,這些基因的表達模式與轉錄組表達模式基本一致,這些基因調控參與了痂囊腔菌素生物合成,為進一步分析提供了研究基礎。











