编译:微科盟 伊一,编辑:微科盟 景行、江舜尧。
微科盟原创微文,欢迎转发转载,转载请注明来源于【转录组】公众号。
导读
由于杨树的连续种植,其生长和生物量积累可能会受到土壤中对羟基苯甲酸(pHBA)等等位化学物质积累的负面影响。光合作用是植物最基本的过程,会受到pHBA胁迫的负面影响。因此,提高pHBA胁迫下的光合作用能力对促进杨树的生长至关重要。丝裂原活化蛋白激酶(MAPK)级联途径广泛参与植物的环境胁迫响应。然而,在pHBA胁迫下,MAPK通路基因对光合作用相关通路的调控机制尚不清楚。 本研究通过转录组分析和加权基因共表达网络分析观察到,PeMPK7在杨树中过表达可调控pHBA胁迫早期光合作用相关基因和转录因子基因(即WRKY1、WRKY33和ERF3)的表达 。此外, PeMPK7还能改善杨树在长期pHBA胁迫下的光合作用 。此外, 酵母双杂交和拉下试验证实了PeMPK7和PeMKK7/10之间的相互作用 。根据这些结果, 本研究构建了PeMPK7参与调控光合作用的途径示意图 。本研究为通过MAPK级联途径调控pHBA胁迫的分子机制提供了新的见解。
论文ID
原名: Potential molecular mechanism of photosynthesis regulation by PeMPK7 in poplar under para-hydroxybenzoic acid stress
译名: 对羟基苯甲酸胁迫下PeMPK7调节杨树光合作用的潜在分子机制
期刊: Ecotoxicology and Environmental Safety
IF: 6.2
发表时间: 2024年4月
通讯作者: 国静
通讯作者单位: 山东农业大学林学院
DOI号: 10.1016/j.ecoenv.2024.116329
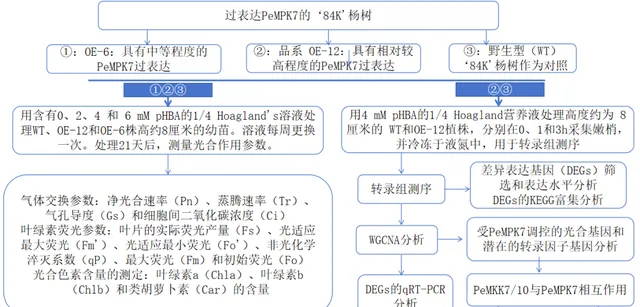
实验设计
结果
1 过表达PeMPK7调控pHBA胁迫下光合作用相关基因的表达
研究者之前的研究报道,PeMPK7能增强杨树对pHBA胁迫的耐受性。 为了研究PeMPK7过表达后杨树早期耐受pHBA胁迫的分子机制,研究者用4mM pHBA处理高度约为8cm的幼苗。1小时后,WT和OE-12植株出现茎尖弯曲和脱水现象 。然而,与WT植物相比,过表达PeMPK7的植物受到的损害要小得多。 处理3小时后,虽然所有植株的茎尖弯曲和叶片脱水程度都有所增加,但WT植株比OE-12植株受到的伤害更大 (补充图S1)。因此, 研究者在对WT和OE-12幼苗施加pHBA胁迫0、1和3小时后,对其叶片进行了转录组测序分析。 这些纯净数据被合并成35422个unigenes,平均长度为1106.93bp,总长度为61984637bp。所有全长转录本都在GO、KEGG、Pfam、Swissprot、eggNOG和NR数据库中进行了注释。
研究者以 p< 0.05和|log2-foldchange|>1作为筛选差异表达基因(DEGs)的标准。 研究者对筛选的DEGs的表达水平进行了分析 (图1A)。 在没有pHBA的情况下,PeMPK7高表达植株(T0)和WT植株(WT0)之间有1659个DEGs(894个上调,765个下调)(T0-vs-WT0) 。 pHBA处理1小时后,PeMPK7基因过表达植株(T1)与WT植株(WT1)之间出现了1671个DEGs(1005个上调,666个下调)(T1-vs-WT1) 。 pHBA处理3小时后,PeMPK7基因高表达植株(T3)与WT植株(WT3)之间出现了1890个DEGs(上调1069个,下调829个)(T3-vsWT3) 。总体而言, 随着pHBA处理时间的延长,PeMPK7高表达植株与WT植株之间的DEGs数量有所增加 。

图1.pHBA处理0、1和3小时后差异表达基因(DEGs)的韦恩图。 (A)PeMPK7过表达(T)和野生型(WT)植株的所有DEGs。(B)T和WT植物光合作用相关DEGs。
研究者在T0-vs-WT0、T1-vs-WT1和T3-vs-WT3对比组中,分别发现了73、52和50个与光合作用相关的DEGs (图1B)。如补充表S2所示, 各对比组分别有66、25和32个基因上调,这些基因主要与采光复合体I/II叶绿素a/结合蛋白、光系统I/II亚基和磷酸布洛激酶有关 。 各对比组分别有7、27和18个基因下调,主要与V型H+转运ATPase亚基C、磷酸甘油酸激酶和光系统I组装蛋白有关 。总体而言, 与WT植株相比,70.9%的光合作用相关基因在过表达PeMPK7的杨树叶片中显著上调 ( p< 0.05)(图2),这些基因主要与光系统I和II亚基以及光合叶绿素结合蛋白有关。因此, 研究者推测过表达PeMPK7的杨树植株比WT植株捕获光能的能力更强。
图2. 在pHBA处理0、1和3小时后,PeMPK7-高表达(T)植株与野生型(WT)植株之间光合作用相关差异表达基因的热图(A)。(B)与光合系统I和II有关的基因。(C)参与卡尔文循环的关键酶。
此外, 研究者还利用KEGG富集分析研究了DEGs参与的主要光合作用途径 。 与光合作用相关的DEGs的两个最显著富集途径是"光合作用(ko00195)"和"光合作用触角蛋白(ko00196) "。此外,在"光合生物的碳固定(ko00710)"等通路中也富集了DEGs(图3B)。
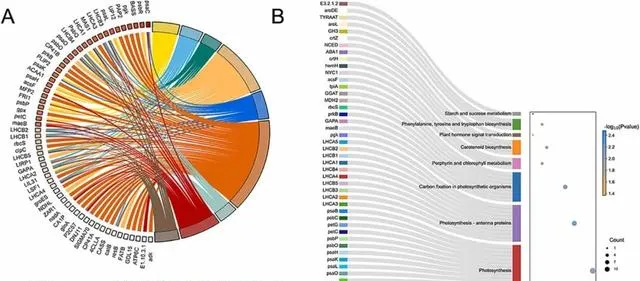
图3.与光合作用相关的差异表达基因(DEGs)的GO和KEGG分析。 (A)共有9个GO术语与DEGs相关。(B)共有8个KEGG通路与DEGs相关。气泡图中圆点的颜色和大小分别代表P值和基因数。
2 WGCNA揭示了光合作用的枢纽基因
研究者根据转录组中的基因,进行了WGCNA分析(图4),共产生了9个共表达模块,每个模块包含93-5880个基因 。在这9个模块中,有5个模块包含超过1000个DEGs,分别是绿松石模块(5880个DEGs)、蓝色模块(2417个DEGs)、棕色模块(2093个DEGs)、灰色模块(3495个DEGs)和黄色模块(1313个DEGs)。 在这些模块中,有三个模块与光合作用的相关性最高,即绿松石模块、棕色模块和蓝色模块,分别有30个、21个和14个光合作用相关基因。因此,研究者选这三个模块作进一步分析的对象。
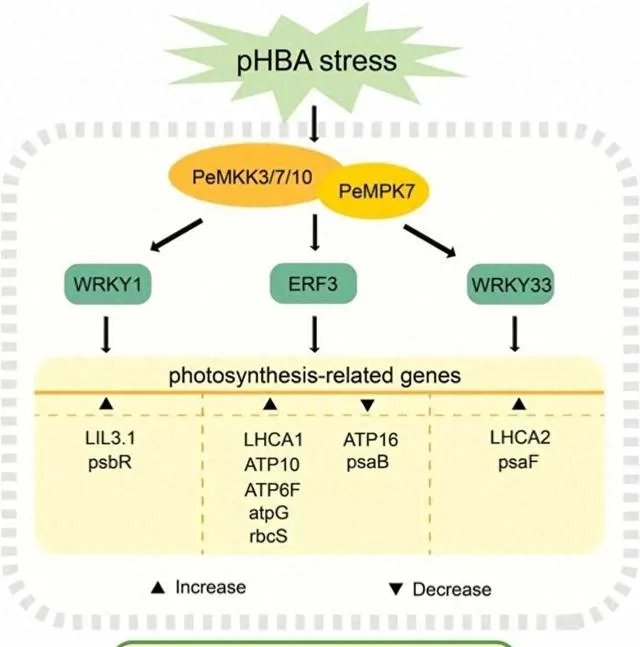
研究者通过构建共表达网络,确定了每个模块中的枢纽基因 。 绿松石色、棕色和蓝色模块分别包括5个(LHCA1、PetA、psaB、psaG和psbK)、4个(ATP6C、psbR、FaQR和LIL3.1)和4个(LHCA2、psbW、psaF和CAAT6)中心基因 (图5C-E)。在上述枢纽基因中, 2个(LHCA1和LHCA2)富集于光合天线蛋白通路,6个(psaB、psaG、psbK、psbR、psbW和psaF)富集于光合作用蛋白通路。研究者推测,这8个与光合作用相关的基因可能有助于在PeMPK7过表达杨树应对pHBA胁迫时维持高水平的光合作用。
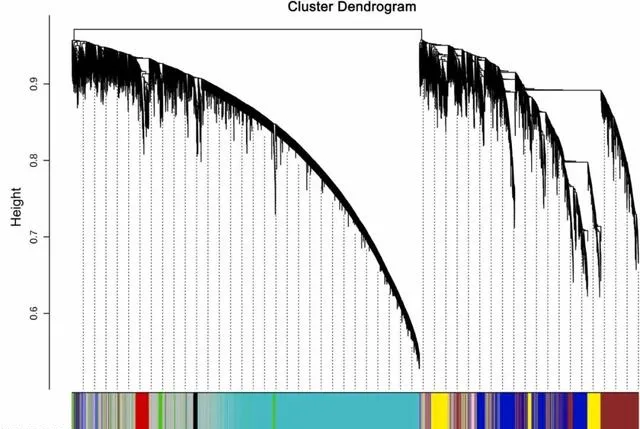
图4.显示加权基因共表达网络识别的共表达模块的层次聚类树。
3 受PeMPK7调控的光合基因和潜在的转录因子基因
研究者根据三个模块的共表达网络,通过分析网络节点关系,构建了光合作用相关子网络 (图5A)。该子网络主要由光合作用相关基因组成,这些基因均为非转录因子(TFs)及其潜在调控TFs。 通过对子网络中的TF进行基因家族分类,研究者发现共表达网络中的光合作用相关基因主要受WRKY(WRKY1和WRKY33)、ZIP(ZIP7和ZTP2)、ERF(ERF3)、bHLH(PIF4)和bZIP(HY5)基因家族的调控 。随着pHBA浓度的增加,HY5、ZIP7、PIF4和ZIP2在PeMPK7过表达植株和WT植株中的表达水平都稳步下降。相反,随着pHBA浓度的增加,WRKY1、WRKY33和ERF3在PeMPK7过表达植株和WT植株中的表达量都有所增加。此外,在pHBA胁迫下,PeMPK7-高表达植株中WRKY1、WRKY33和ERF3的表达水平显著高于WT植株(图5B)。 研究者推测PeMPK7可能通过调节WRKY1、WRKY33和ERF3等TFs的表达来积极调节杨树在pHBA胁迫下的光合作用。
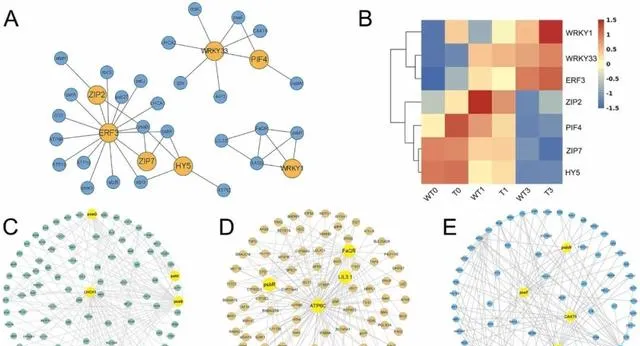
图5.差异表达基因的共表达网络分析。 (A)与光合作用相关的子网络。橙色圆圈代表转录因子(TFs),蓝色圆圈代表与光合作用相关的基因。(B)热图显示转基因植物和WT植物在三个时间点的TFs表达水平。(C-E)黄色圆圈代表枢纽基因。
4 DEGs的qRT-PCR分析
为了验证转录组的结果,研究者从转录组数据中随机选择了5个与光合作用相关的DEGs(LHCB1、LHCB5、PsaC、PsbR和LIL3.1)和6个其他DEGs(GALE、PP2C、SPG20、PIF4、rpiA和AUX1)进行qRT-PCR分析 。在pHBA处理后0、1和3小时,PeMPK7过表达植株中LHCB1、LHCB5、PsaC、PsbR、LIL3.1和AUX1的表达水平均高于WT植株。在pHBA处理3h后,PeMPK7基因表达植株中GALE、PIF4、PP2C和SPG20的表达水平高于WT植株,而rpiA的表达水平低于WT植株(图6)。 这11个基因的相对表达水平与转录组结果一致,证实了转录组数据的准确性 。此外,这些结果表明过表达PeMPK7可提高光合作用相关基因如LHCB1、LHCB5、PsaC、PsbR和LIL3.1的表达水平,使植物在pHBA胁迫下保持较高的光合速率。
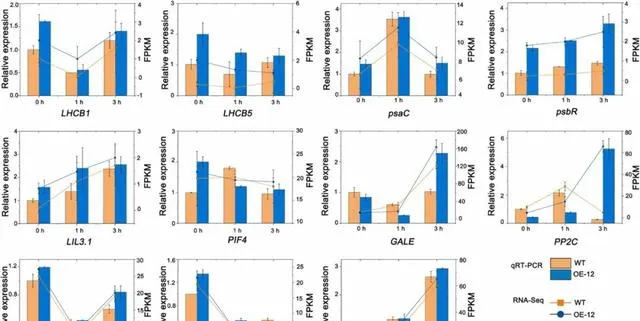
图6.转录组中差异表达基因的qRT-PCR验证。数据以三个重复的平均值±SD表示。
5 过表达PeMPK7可提高pHBA胁迫下杨树的光合作用
为了进一步研究PeMPK7是否参与调节杨树在长期pHBA胁迫下的光合作用,研究者对OE-6、OE-12和WT植物分别用0、2、4和6mM pHBA处理。在pHBA处理的第21天,测量植株的光合特性。
pHBA处理前,OE-6和OE-12中Chla、Chlb和Car的含量显著高于WT植株,但OE-6中Chlb的含量除外( p< 0.05)。 经pHBA处理后,三种植物中Chla、Chlb和Car的含量均以浓度依赖性的方式下降 。 经6mM pHBA处理后,三种植物中Chla、Chlb和Car的含量最低 。然而, OE-12和OE-6植株中的含量高于WT植株 (图7)。与暴露于0mM pHBA的植株相比,OE-6、OE-12和WT植株的Chla含量分别降低了21.15%、27.68%和38.83%。因此, 研究者认为pHBA处理抑制了杨树叶片中叶绿素的生物合成或加速了叶绿素的降解 。然而, 在杨树中过表达PeMPK7可促进Chla、Chlb和Car的合成或缓解它们的降解。
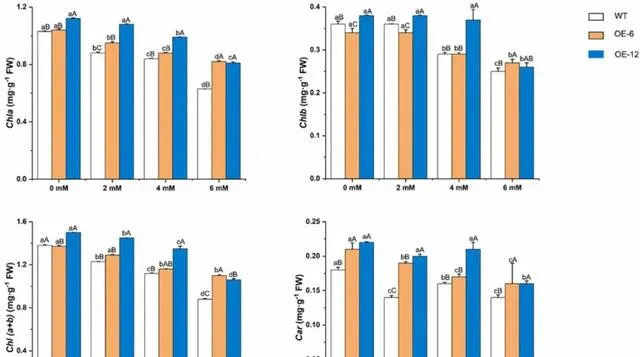
图7.pHBA胁迫对过表达PeMPK7植物叶片光合色素含量的影响。 数据以三个重复的平均值±SD表示。不同字母表示差异显著( p< 0.05)。
在pHBA胁迫处理前,OE-6和OE-12植株的Pn显著高于WT植株( p< 0.05);但在Tr、Gs、Ci和Ls方面,OE-6、OE-12和WT植株没有观察到显著差异。 经pHBA处理后,随着pBHA浓度的增加,三种植株的Pn、Tr、Gs和Ls均下降,Ci上升。 经2mM pHBA处理后,OE-12和OE-6植株的Gs和Ls显著高于WT植株( p< 0.05)。用4mM pHBA处理后,OE-12和OE-6植株的Ls明显高于WT植株。经6mM pHBA处理后,OE-12和OE-6植株的Pn、Tr、Gs和Ls显著高于WT植株( p< 0.05),Ci显著低于WT植株(图8)。 这些结果表明,过表达PeMPK7能改善高浓度pHBA条件下杨树的Pn和蒸腾作用,并且过表达PeMPK7的杨树叶片的气孔和Ci的调节能力优于WT植株。
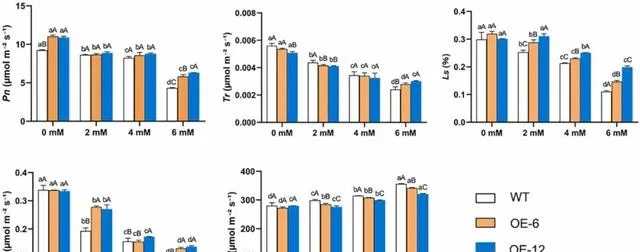
图8.pHBA胁迫对过表达PeMPK7植物叶片气体交换参数的影响。 数据以三个重复的平均值±SD表示。不同字母表示差异显著( p< 0.05)。
pHBA处理前,OE-6、OE-12和WT植株的Fv/Fm、Fv/Fo、ΦPSII、qP和NPQ无明显差异。pHBA处理后,当pHBA浓度低于6mM时,Fv/Fm、Fv/Fo、ΦPSII和qP的值随pHBA浓度的增加而降低,而NPQ则随pHBA浓度的增加而升高 。经6mM pHBA处理后,三种植株的Fv/Fm、Fv/Fo、ΦPSII和qP值均最小,OE-12和OE-6植株的Fv/Fm、Fv/Fo、ΦPSII、qP和NPQ值均高于WT植株(图9)。 这些结果表明,pHBA胁迫显著抑制了杨树叶片的光合效率、PSII电位活性、ΦPSII和电子传递。 随着 pHBA浓度的增加,OE-12和OE-6植株的光化学效率、电子传递速率、PSII反应中心开放度和调节能力均明显高于WT植株。
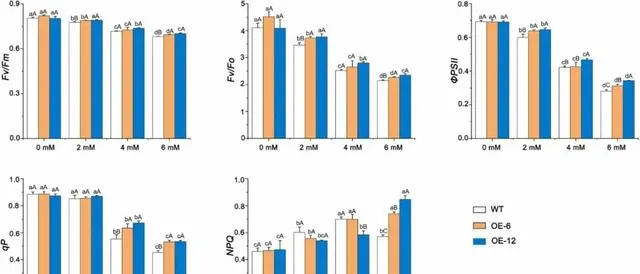
图9.pHBA胁迫对过表达PeMPK7植物叶片叶绿素荧光参数的影响。 数据以三个重复的平均值±SD表示。不同字母表示差异显著( p <0.05)。
6 PeMKK7/10与PeMPK7相互作用
MAPK在被其上游MPKK磷酸化后参与植物防御反应。研究者之前的研究报道了PeMPK7与PeMKK3之间的相互作用。 为了进一步了解PeMPK7在调控杨树对pHBA胁迫响应中作用的分子机制,研究者对PeMPK7与其他MPKKs的相互作用进行了研究。 根据STRING数据库的预测,PeMKK3、PeMKK7、PeMKK9和PeMKK10的得分高于其他MPKKs。因此, 研究者进行了酵母双杂交和拉下试验来分析PeMPK7和PeMKK7/9/10之间的蛋白质相互作用。
在酵母双杂交试验中,PeMPK7与pGBKT7载体中的GAL4DNA结合域融合,而PeMKK7、PeMKK9和PeMKK10则克隆到pGADT7载体中的GAL4激活域 。结果表明, 阳性对照、阴性对照和pGBKT7-PeMPK7+pGADT7-PeMKK7/9/10共转化的酵母菌株在SD/-Trp/-Leu固体培养基上生长正常 。除阴性对照外,所有菌株均在SD/-Trp/-Leu/-Ade/-His/X-α-Gal/100ng/mLAbA固体培养基上生长,并在X-α-Gal染色后呈现着色(图10A)。 这些结果表明,PeMPK7可能与PeMKK7、PeMKK9和PeMKK10相互作用 。 为了进一步证实这一点,研究者使用融合蛋白PeMPK7-His和PeMKK7/9/10-GST进行了拉下试验 。结果表明 ,在体外,PeMPK7-His与PeMKK7-GST和PeMKK10-GST形成了异二聚体,这表明PeMPK7与PeMKK7/10相互作用,而不是与PeMKK9相互作用 (图10B-D)。
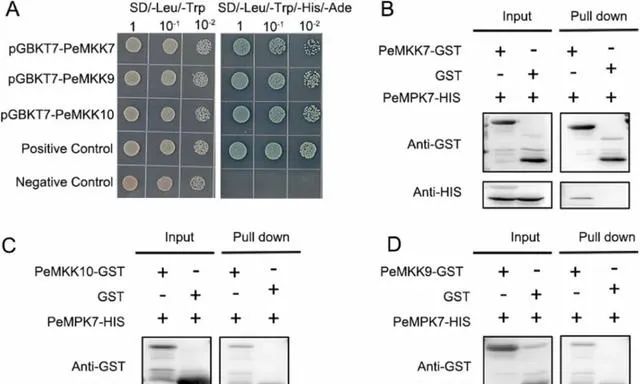
图10.PeMPK7与PeMKK7/10相互作用。 (A)酵母双杂交试验。菌株在SD/-Trp/-Leu和SD/-Trp/-Leu/-Ade/-His/X-α-Gal/AbA平板上生长的能力表示各自的蛋白-蛋白相互作用;1、10-1和10-2代表酵母稀释度。pGBKT7-53+pGADT7-T和pGBKT7-Lam+pGADT7-T的酵母菌株分别作为阳性对照和阴性对照。(B-D)为验证PeMPK7和PeMKK7/9/10之间的蛋白相互作用,进行了牵引试验。在大肠杆菌中诱导PeMPK7-GST和PeMPKKs蛋白,空GST蛋白作为对照。
7 PeMKK7/10的表达受pHBA调控
为了研究PeMKK7/10的表达是否受pHBA的调控,研究者对经pHBA处理的WT植株进行了qRT-PCR。经pHBA处理1、3和6h后,PeMKK7和PeMKK10的表达均被显著诱导。
讨论
光合作用是植物交换气体和合成能量的主要方式。环境因素会影响植物的光合特性。pHBA胁迫会破坏叶片中叶绿体的结构和功能,抑制光合作用,导致植物无法维持正常的生理活动。 本研究通过转录组和光合生理指标分析证明,PeMPK7过表达可提高杨树应对pHBA胁迫的光合作用。
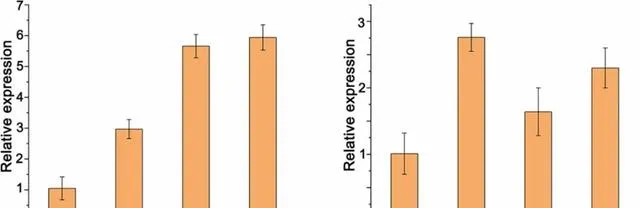
图11.pHBA处理下杨树中PeMKK7和PeMKK10的表达分析。 数据以三个重复的平均值±SD表示。
1 PeMPK7参与了pHBA胁迫下杨树叶绿素和类胡萝卜素的合成和代谢
叶片是植物的主要光合器官,对作物的品质和产量至关重要。叶绿素浓度以及叶绿体的形状和结构直接影响光合作用的效率。叶绿素、碳水化合物、脂质和蛋白质复合物构成了光合系统,这是一个吸收光的功能单元。通过吸收和传递能量,叶绿素在光合系统中发挥着特殊而关键的功能。此外,叶绿素的生物合成、积累和降解还与叶绿体发育、光形态形成和叶绿体-细胞核信号传递有关。PSI和PSII中的主要色素是Chla和Chlb,它们在Lhc蛋白中捕获光能。Car在光保护中起着至关重要的作用。在外源FA处理下,草莓叶片中Chla、Chlb和Car的含量下降,导致光合作用速率降低。之前的研究报道,SlMAPK3的过表达增加了番茄的叶绿素含量和光合作用活性。与这些研究结果一致, 本研究表明,在pHBA胁迫下,杨树叶片中Chla、Chlb和Car的含量显著下降 (图7)。此外, 在高浓度pHBA胁迫下,过表达PeMPK7会促进杨树叶片中Chla、Chlb和Car的积累。同样,PeMPK7的过表达也影响了参与血红素和叶绿素代谢以及Car生物合成、运输和降解过程的基因的转录水平 。 在本研究中,与血红素和叶绿素代谢相关的DEGs在过表达PeMPK7的植株中显著上调 ,这与Ma等人的研究一致。 在本研究中,研究者通过WGCNA和共表达网络构建分析确定(图5D)采光复合物样蛋白3同型1(LIL3.1)为棕色模块中的一个枢纽基因。 采光复合物蛋白可以正向调节叶绿素的生物合成,在光合作用中发挥关键作用。 qRT-PCR研究结果表明,在pHBA处理0、1和3小时后,PeMPK7过表达植株的LIL3.1转录水平高于WT植株 (图6)。因此,在pHBA胁迫下,PeMPK7可通过调节叶绿素的合成和代谢以及碳水化合物的生物合成和降解过程来维持转基因植物的正常光合作用。
2 PeMPK7参与了pHBA胁迫下杨树的光合碳循环过程
光合效率下降是光抑制的最显著特征,它反映了光合系统的运行状况。酚类化合物可通过影响膜电位、渗透性和活性来影响细胞膜功能。它们将信号从根部传递到地上部分,影响植物体内的酶活性和渗透胁迫,并导致气孔关闭,从而导致植物光合速率下降。pHBA能显著抑制杨树的光合作用,且抑制作用随pHBA浓度的增加而增强,其主要原因是非气孔限制。本 研究结果表明,随着pHBA浓度的增加,杨树叶片的Ci增加;Ls显著降低,Pn逐渐降低 (图8)。这与之前关于酚酸胁迫下杨树Pn和Ls下降的研究结果一致。 这表明非气孔限制是促进Pn下降的主要因素;这可能是由于在pHBA胁迫下叶绿体和类叶绿体结构被破坏,导致光合活动紊乱和叶绿素含量下降 。然而, OE-6和OE-12植株Pn的下降幅度明显低于WT植株,说明PeMPK7的过表达能有效提高植株的净光合效率 。此外, 随着pHBA浓度的增加,杨树的Tr和Gs逐渐降低。这表明,pHBA胁迫会导致杨树气孔关闭、蒸腾作用减弱、水分代谢能力下降 。 在高浓度pHBA胁迫下,PeMPK7表达植株Tr的降低程度明显低于WT植株,说明在高浓度pHBA胁迫下,PeMPK7表达植株的水分代谢能力优于WT植株 。之前有研究报道,在杨树中过表达PeNAC122能在渗透胁迫下维持较高的气体交换速率。 转录组分析表明,在pHBA胁迫下,PeMAPK7过表达植株中NAC家族基因如NAC104、NAC55、NAC2和NAC72的表达水平显著高于WT植株。 因此, 研究者推测PeMPK7通过调节这些NAC家族DEGs来维持较高的气体交换率 。这一假设需要通过分子生物学实验进一步验证。转基因水稻中Rubisco含量的增加和Rubisco活性的维持可在最适温度范围内的中等高温条件下提高光合作用。同样, 转录组分析表明,与WT植株相比,PeMPK7的过表达显著上调了植物卡尔文循环中编码酶的大多数关键基因,如RbcS、maeB、cynT、can、pgk、prkB和GAPA (图2C和12)。 研究者推测PeMPK7的过表达可能会显著提高Rubisco的羧化和RuBP的再生,从而提高植物的光合能力 。结合之前关于PeMPK7改善渗透调节能力的研究, 研究者推测PeMPK7可能会增加渗透调节物质的含量,降低渗透势,维持细胞的张力压力,促进植物体内各种物质代谢的顺利进行,从而维持较高的光合效率和蒸腾速率 ,这有助于植物抵抗pHBA胁迫。
3 PeMPK7参与了pHBA胁迫下杨树光能的吸收和转化
叶绿素荧光参数作为植物适应各种环境的内在指标,可用于确定环境胁迫下光合器官的受损程度。例如,Fv/Fm是PSII的最大光化学效率,反映了反应中心的初级光能转换效率;qP反映了PSII触角色素吸收的光能中用于光化学电子传递的份额;ФPSII反映了PSII反应中心的实际光能捕获效率。在pHBA胁迫下, Populus × euramericana ‘Neva’和Rumex acetosa的Fv/Fm和ФPSII都有所下降。同样, 在本研究中,pHBA胁迫后杨树的Fv/Fm、ФPSII和qP显著降低。这些结果表明,pHBA胁迫会对光合器官造成损伤。
有趣的是, PeMPK7过表达植株的Fv/Fm远大于WT植株 (图9), 且在4mM和6mM pHBA处理后,OE-6和OE-12植株ФPSII和qP的下降幅度远小于WT植株 (图9)。 这表明过表达PeMPK7可增加光化学转移所需的光能,从而提高pHBA胁迫下植物光合电子传递的效率 。与研究者的结果一致,在干旱胁迫下,过表达SlMAPK3的番茄植株的ФPSII和qP值均高于WT植株。由于SlMAPK3和PeMPK7分别属于A组和C组MAPK, 研究者推测A组和C组MAPK基因在非生物胁迫下具有提高光合电子传递效率的保守功能。
在锌胁迫下,SbMT-2转基因烟草植株的qP值和ФPSII值均高于WT植株,这表明SbMT-2的过表达可以提高植物光合电子传递的效率。 在本研究中,转录组分析表明,在pHBA胁迫下,PeMPK7基因过表达植株表达的MT-2水平大大高于WT植株。 PeMPK7可能在MT-2基因的上游发挥作用,并通过调节MT-2的表达提高杨树的光合效率。这一点应在今后的研究中加以验证。入射光、环境温度、微量元素和水分供应的变化对光合作用有很大影响。当入射光水平超过光合作用的吸收能力时,会对植物造成损害,导致光损伤。为了避免光损伤,植物会利用NPQ机制将多余的能量以热量的形式耗散。NPQ有多种类型,如qE、qH、qZ和qL。qE机制依赖于类囊体蛋白PsbS和黄绿素的可逆光依赖性转化。受到光照后,质子梯度会在类囊体膜上形成。这引发了PsbS的质子化,并通过激活中黄素脱氧化酶等酶产生玉米黄质,然后将其淬灭。 本研究结果表明,当pHBA浓度低于6mM时,过表达PeMPK7和WT杨树叶片的NPQ随pHBA浓度的增加而增加。然而,当pHBA浓度为6mM时,WT植物的NPQ迅速下降,而PeMPK7基因表达植物的NPQ仍呈上升趋势 (图9)。这一结果与Li等人的研究结果不一致。 研究者推测,当pHBA浓度高于6mM时,超过了WT杨树的调节范围,对PSII反应中心的光合装置造成严重破坏。这导致了PSII反应中心的破坏。结合转录组分析结果,PeMPK7过表达植株的PsbS表达量明显高于WT植株( p <0.05) (图2B和图12)。 这表明过表达PeMPK7能增加NPQ和散热,保护PSII反应中心的光合装置,并通过调节PsbS的表达维持其正常的光合作用 。这与之前利用VIGS技术在牡丹中瞬时沉默PlMAPK1的研究结果一致;该研究报道转基因植株的NPQ低于WT植株。因此,研究者推测C组中的MAPK1和MAPK7基因可能在改善NPQ和增加热耗散以保护PSII反应中心的光合装置和维持光合作用方面具有功能保护作用。
在本研究中,转录组分析显示PSI和PSII系统中的LHCA1-4、LHCB1-5、petC、psaC、psaK、psaO、psaH、psaL、psbP、psbO和psbR以及Cytb6f中的PetC等基因上调 (图2A、2B和12)。同时, WGCNA和共表达网络的构建确定了LHCA1、LHCA2、PetA和FaQR基因是光合作用过程中的枢纽基因 (图5C-E)。 这些结果表明,过表达PeMPK7可通过增强光能吸收和光合电子传递能力来提高光合效率。
4 PeMPK7参与pHBA胁迫下光合作用相关基因的TF介导调控
TFs通过与目标基因启动子中的顺式调控元件结合来调控基因表达。鉴于多个光合作用相关基因在转录本水平上受到pHBA的调控,研究者认为TFs在pHBA介导的这些基因的表达中发挥着重要作用。 本研究通过构建与光合作用相关的子网络,揭示了TFs与光合作用相关基因之间的潜在调控关系 (图5A)。
本研究构建的光合作用相关子网络主要包括编码采光叶绿素(LHC)a/b和PSI/II反应中心色素蛋白的基因以及具有调控关系的潜在TFs。 潜在的TFs属于WRKY、ZIP、ERF、bHLH和bZIP基因家族。据报道,这些TF的同源物参与了光合作用的调控。例如,植物色素相互作用因子(PIFs)是叶绿素和Car生物合成的负调控因子,植物色素部分抑制PIFs以实现光自养生长。bZIP成员HY5是一种有效的PIF拮抗剂,它能促进光合色素的积累以应对光照。此外,HY5还能控制参与光合作用的其他基因的转录,包括核酮糖-1,5-二磷酸羧化酶/氧合酶(RbcS1A)的小亚基。此外,非典型bHLH TF SlPRE5负向调节转基因番茄叶片中的叶绿素含量,PebHLp5通过调节光合作用正向调节干旱胁迫响应。因此,光合作用相关子网络中预测的潜在IFs可能参与调控光合作用相关基因。
迄今为止,尚不清楚植物MAPK基因利用哪些TFs来调控pHBA胁迫下的光合作用。 研究者对PeMPK7过表达植株和WT植株中TFs表达水平差异的分析表明,光合作用相关子网络中的ERF3、WRKY1和WRKY33(图5A)在PeMPK7过表达植株和WT植株中都随着pHBA浓度的增加而上调 。此外, 在pHBA处理1和3h时,这三个基因在PeMPK7高表达植株中的表达水平明显高于WT植株(图5B)。这表明在pHBA胁迫下,PeMPK7通过正向调节WRKY1、WRKY33和ERF3的表达参与光合作用的调控。 在盐胁迫下,转基因菊花中的DgWRKY5可通过增加Pn、Gs和Tr来改善光合作用和生长。ERF蛋白(ERE-结合蛋白,ERE BPs)是从烟草中分离出来的GCC盒结合蛋白。先前的研究利用拟南芥蛋白质微阵列确定了拟南芥中作为MAPKs底物的15个ERF家族成员。其中,拟南芥中病原体反应的正调控因子AtERF104通过与MPK6相互作用而稳定。水稻中的OsEREBP1基因可被BWMK1(病原体诱导的MAPK)磷酸化,从而增加其在体外与顺式元素的DNA结合活性。这些研究提示了PeMPK7和ERF3之间潜在的蛋白质相互作用。在pHBA胁迫下,PeMPK7通过正向调节WRKY1、WRKY33和ERF3来调控光合作用,这一点还需要利用遗传学和分子生物学方法进一步验证。
5 PeMPK7参与了响应pHBA应激的级联信号通路
参与植物抗逆的MAPK级联对于了解MAPK级联途径在响应pHBA胁迫时的调控分子机制至关重要。据报道,有几种MAPK信号通路参与了植物对生物和非生物胁迫的响应。例如,水稻中的OsMEKK6-OsMAPK5-SERF1信号通路参与了对盐胁迫的响应,拟南芥中的AtMEKK1-AtMKK1/AtMKK2-AtMPK4信号通路参与了对干旱胁迫和机械损伤的响应。然而,有关MAPK信号通路参与杨树胁迫响应调控的研究却很少。过表达PtMKK4能显著增强杨树的抗旱性,并提高过表达PtMKK4的杨树中PtP5CS、PtSUS3、PtLTP3和PtDREB8的表达水平。研究者之前的研究证明了PeMPK7和PeMKK3之间的相互作用。 本研究表明,除PeMKK3外,PeMPK7还与PeMKK7/10相互作用(图10),并且PeMKK7和PeMKK10的表达受pHBA的调控。因此,研究者推测PeMKK3/7/10-PeMPK7通路是参与杨树对pHBA胁迫响应的重要MAPK信号通路 。然而,这三条途径的参与还需要通过体外磷酸化实验进一步验证。此外, 研究者的转录组结果显示PeMPK7可能调控下游TFs,即WRKY1、WRKY33和ERF3,这表明PeMKK3/7/10-PeMPK7通路可能通过直接或间接调控WRKY1/WRKY33/ERF3参与了对pHBA胁迫的响应 (图12)。 鉴于WRKY1/WRKY33/ERF3可能调控光合作用相关基因(图5A和12),研究者推测PeMKK3/7/10-PeMPK7模块通过调控WRKY1/WRKY33/ERF3来调控光合作用相关基因。 这一假说有待今后利用遗传学和分子生物学技术进一步验证。
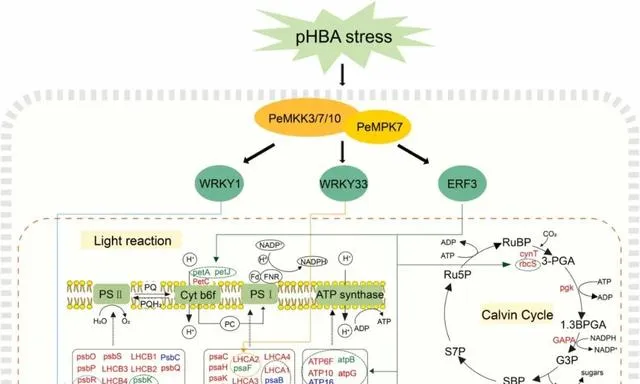
图12.光合作用相关途径示意图。 红色、蓝色和绿色表示与WT植物相比,PeMPK7基因过表达植物在pHBA胁迫后显著上调、下调和无显著变化的基因。
结论
总之,
PeMPK7在杨树中的过表达即使在pHBA胁迫下也能维持高水平的光合作用。本研究通过转录组分析、WGCNA和光合作用相关指标的测定,探讨了PeMPK7在pHBA胁迫下调控光合作用的分子机制
。
PeMPK7在杨树中的过表达能在pHBA胁迫早期增加光合作用相关基因的表达。WRKY、ZIP、ERF、bHLH和bZIP基因家族的TFs可能与光合作用相关基因的调控有关。PeMPK7可能在WRKY1、WRKY33和ERF3的上游发挥作用,并可能通过正向调节它们的表达来促进杨树的光合作用。
此外,
光合生理指标分析证实,在长期pHBA胁迫下,PeMPK7参与了杨树光合作用的调控
。此外,
研究者利用酵母双杂交和拉下实验证明了PeMPK7和PeMKK7/10之间的相互作用。基于这些结果,研究者推测PeMKK3/7/10-PeMPK7-WRKY1/WRKY33/ERF3通路是参与杨树对pHBA胁迫响应的重要潜在MAPK信号通路
。通过构建PeMPK7调控光合作用的通路模型,研究者对MAPK级联通路调控pHBA胁迫的分子机制有了新的认识。











