文 | 追問nextquestion,作者 | 十三維,編輯 | 存源
生命是一個復雜而神秘的現象。費曼曾說:「What I can't create, I do not understand」。如果要理解生命,最好的方式恐怕就是去創造生命。無論是古猶太的魔偶(Golem)、古希臘的塔羅斯、中國周朝的偃師,還是文學作品中的法蘭克史坦和現代科幻小說,這種創造生命的沖動跨越不同文明,貫徹整體人類歷史。而今天,這種創造生命的沖動不單是為了滿足功能或生產力的需求,更在科技實踐中展現其潛力,尤其是在人工生命和人工智慧領域,為理解自身和我們在宇宙中存在的意義做出了重要貢獻。
人工生命(Artificial Life, Alife)是一個新興的跨學科研究領域,它致力於透過計算模型和物理模擬,探索生命的各種潛在形式。 Alife不僅試圖模擬和復制地球上已知的生命形式,還嘗試創造全新的、可能存在的生命形式,從而擴充套件我們對「生命」這一概念的理解。 這包括使用電腦模擬生命過程,研究自組織系統,以及開發能模擬自然選擇和前進演化過程的軟體和硬體系統。本文將在回顧Alife歷史的基礎上,介紹其最新的研究方向和實踐進展。
從生命到人工生命
傳統生物學中,生命通常被認為是由DNA和蛋白質構成,具有新陳代謝、生殖發育、遺傳演化等一系列性狀的自然系統。直到1944年,物理學家薛丁格發表【生命是什麽】( What Is Life? ),生命才擺脫特定的生理組織結構依賴,被定義為一個能量-資訊耦合的負熵系統。這種思想既回應了過去,也迎合了當下。將生命從純粹的有機論拉回到了有機論和機械論之間,也為Alife的理論研究提供了可能。
就在同一時期,許多不同領域的科學家開始了對Alife的探索,這一領域的探索大致可以分為三次浪潮:
第一次(20世紀50-60年代):自復制探索期
在這個時期,科學家們主要關註自復制、自組織和前進演化等生命維持和產生結構的基本原理。涉及的領域包括控制論、圖靈機、形態發生學、神經網路模型和遺傳演算法。尤其是1948年馮·諾依曼發表【自復制自動機理論】,奠定了人工生命學科基礎。
這一次浪潮, 科學家們將生命視為一種邏輯形式,主要強調資訊處理的抽象模型。 在實踐上,挪威數學家尼爾斯·貝瑞切利(Nils Barricelli)在1953-1962年於早期的電腦IAS上,便使用了一維元胞自動機,模擬了一個可以無限前進演化的數位世界。其中甚至出現了數位共生體生命形式。他據此認為,基於競爭前進演化模型的達爾文理論不足以解釋生命演化的全貌,共生和合作在生命演化中占有重要地位 [1] 。這次浪潮從模型和演算法層面梳理了生命的基本特征,祛除了生命發生的迷思,為人工生命奠定了理論和實踐基礎。
第二波(20世紀70-90年代):計算模擬期
這一時期, 科學家們開始大量利用計算模擬技術,包括元胞自動機、神經網路、數位前進演化系統等,來研究適應力、湧現等生命系統特性。
1987年9月,凱瑞斯托弗·蘭頓(Christopher Langton)組織了第一次人工生命會議,這標誌著人工生命學科的誕生。會議中, ALife被定義為對「可能存在的生命」 (life as it could be) [2] 的研究, 與僅限於地球已知生命形式的傳統生物學形成對比。其中馮·諾依曼的元胞自動機,隨後被約翰·康威(生命遊戲)、史帝芬·沃弗拉姆,以及蘭頓本人等發展,成為Alife最重要的理論模型。
蘭頓定義了一個元胞自動機活性參數λ,當λ適中時,元胞自動機會兼具局域化的穩定模式和無組織混沌行為,即「混沌邊緣」的自動機。他認為生命或智慧均起源於混沌邊緣。這也是復雜系統理論的重要思想。第二次浪潮,在發展模型基礎上開始大量引入計算模擬技術,從實踐上推動研究生命的適應力和湧現等復雜系統特征,標誌了人工生命學科的建立和工程實踐的開始。
前兩次Alife所持的功能主義立場以及抽象模型模擬,有助於抓住生命普遍本質特征。正如蘭頓所相信, 生命是一種獨立於載體的恰當形式,不同的物質可以實作相同的生命和智慧功能 [3] 。因此,這兩次浪潮下的主要產物是軟體/虛擬/數位人工生命(Soft)。

然而,在Alife研究的發展過程中,質疑和反思始終伴隨其左右。同期活躍的理論生物學家勞勃·羅森(Robert Rosen),在1991年出版了代表作【生命本身】( Life Itself ),認為生命體是一個「有效因果閉合」(closed to efficient causation)的(M,R)系統* [4] :即系統中驅動變化發生的"因",本身就必須是這個系統的"果"。例如生命系統中酶就扮演了有效因果閉合的角色,它們催化代謝反應的發生又產生自身。這種迴圈因果結構使生命具有某種自主性和自生成能力,意味著生物體不完全是機器,無法完全透過圖靈機模擬。
(M,R)系統:即代謝(Metabolism)和修復(Repair)。代謝由一組機制(記為f)完成從材料A到產物B的轉化,修復則由一組機制(記為Φ)從代謝產物B合成代謝機制f本身。
與此同時,生物和認知科學家法蘭西斯科·瓦雷拉(Francisco Varela)也反對蘭頓的觀點 [5] 。他認為生物體的情境性和歷史性不可還原,生物體與其環境的具身互動遠比其潛在的邏輯或功能模式更重要。這些思考引出了第三次人工生命浪潮。
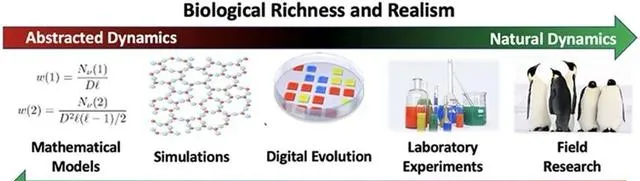
▷ Alife研究方法光譜:抽象模型、模擬仿真、數位前進演化、生化實驗、實地研究,體現了從抽象模型到現實嵌入、以及軟、硬、濕、混合範式的發展歷程。 [6]
第三波(20世紀90年代後期至今):嵌入式演化期
這一時期的研究更加關註人工生命與現實環境的關系, 強調生命在真實世界的嵌入性、具身性、互動性和湧現性。 1972年和1991年,瓦雷拉分別和合作者提出自創生(Autopoiesis)和具身心智(Embodied Mind)思想,分別強調生命系統的自我生成能力,以及與環境具身互動而產生的動態、嵌入式的湧現過程。這些都為人工生命研究提供了新的理論視角。
例如,研究者試圖在系統內部引入「向上的張力」(避免系統落入平衡態的能力和結構),構建自指演化和動力學 [7] ,以及將環境和觀察者(如註意力和操作)納入到系統功能描述中;或者在實驗室進行實驗,推動其在自組織遞迴迴圈中不斷突破和產生創新功能和行為,在一個生態系中向更廣闊的方向演化。
除了數位人工生命外,濕件(Wet)、硬體(Hard),以及混合人工生命(Hybrid),成為這一時期的代表。這包括分子-細胞層面的人工化學、合成生物學、自復制機器人等領域的探索, 研究範圍從人工生命物發展到人工生態系和社會 。即便是在數位人工生命的研究中,科學家的重點也放在數位具身形式 [8] 和數位生態上。

▷ EvoSphere:自主演化機器人系統,在環境中不斷選擇收集核廢料能力個體,再透過無線「交配」最佳化基因型,透過3D打印技術產生新的機器人後代。 [9]
總得來看,我們可以把Alife發展分為兩個軸,橫軸是技術實作的方式,從軟體計算模擬(軟)→物理硬體實作(硬)→生物化學實驗(濕)→仿生混合系統(混合),縱軸是研究焦點,從人工生物體 → 人工生態 → 開放式演化 → 共生(如下圖)。
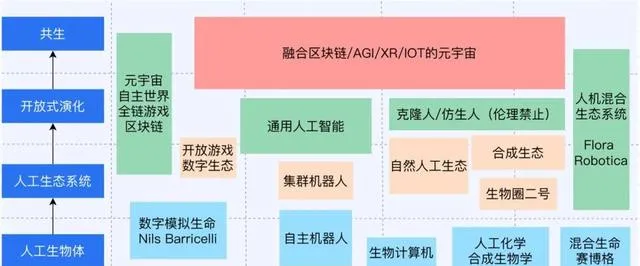
人工生態與社會
在自然界中,生物通常不是孤立存在的,例如,許多靈長類動物和社會性昆蟲以族群的形式在生態系中共生。生態系,作為推動生命演化的核心力量,也由環境及其中的所有生物構成,表現為一個充滿生態互動的動態系統。生態學的研究可以分為群落生態學和生態系生態學,這兩者分別關註生物族群間的互動及生物與物理環境之間的互動。與單純創造人工生物體不同,這一分支的Alife研究的是交互作用的人工實體的集合。
在自然生態群落中,不同物種的生物交互作用。在Alife中,研究者則經常將個體視為某種基因型、表現型或生態型(genotypes,phenotypes,ecotypes)來分析群落動態。威廉·萊納斯(William Reiners) [10] 提出,統一的系統生態學研究需要至少三個獨立互補的理論框架:能量學、物質學,以及族群交互作用或生態系「環通度」(connectedness)的演化。這些框架可以分別理解為能量流、物質流和因果資訊。
其中能量-物質的流動和轉換,包括營養級、食物鏈和網 [11] 、生產率,地球生化學迴圈(如碳迴圈)等;族群互動或生態系的環通度則包括族群之內的集體行為 [12] 、群落之間的交互作用(如捕食、競爭和共生)以及對生態席位的研究,探索這些生態席位如何湧現,消失,然後前進演化的 [13] 。
對ALife人工生態演化的研究,理論技術主要包括復雜動力系統、元胞自動機、反饋網路和控制論等,數位前進演化模擬平台則提供了檢查復雜前進演化和生態系動態的工具。
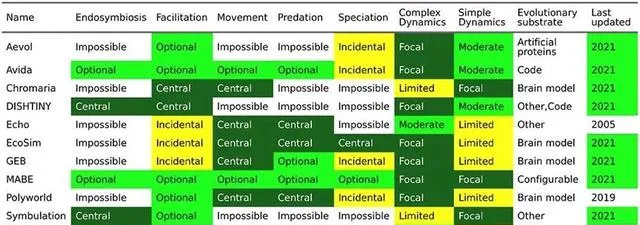
▷ 常用Alife數位前進演化模擬平台。 [6]
其中一些模擬平台設定了能量-物質流模擬。例如,在Tierra中,能量被定義為執行指令所需的CPU周期(電腦處理時間)。智慧體消耗CPU周期執行指令,改變其局部環境。因此,最小化能量消耗(CPU使用)可以提高復制效率,這為智慧體施加了選擇壓力,促使其朝向更高效、可靠的演化方向發展。而在Avida平台中,生物體則透過代謝不同的有限資源(能量)相互競爭。
ALife 生態研究系統與自然系統不同之處在於,研究者可以設定遠離自然界常見值的參數 [15] 。這樣的設定就使得研究者能夠探索哪些特征是生命所必需的,哪些特征和事件可能只是地球上生命形式的偶然產物。這樣,ALife不僅能「重放前世的磁帶」,還可以設計和播放與地球已存或可能執行非常不同的磁帶(如火星),從而幫助澄清和提取生命的本質內容。
人工生態構建也分為軟、硬、濕及混合人工生態四種形式。Alife數位平台能天然模擬軟人工生態,其中還包括人工生命軟體系統或遊戲、及基於前進演化計算的數位生態藝術等。硬人工生命生態主要涉及機器集群系統的互動。濕人工生命生態則嘗試構建通常不會共存的生物組成的生態群落,如執行理想任務(分解廢物、封存碳等)的生態群落,這個子領域被稱為合成生態學。至於混合人工生態,最著名的是 Flora Robotica 計畫 [16] ,該計畫建立了一個植物和機器人共生的生態系,機器人會透過在不同位置發射不同顏色的光來控制植物生長。

▷ 軟體、濕件、硬體人工生態系。
人工生態系領域的研究和實踐非常廣闊。例如,隨著復雜性的增加,生態系可以促進社會和文化互動 [17] 。其中涉及的掠奪、合作和文化的互動,引發了對感知、交流和語言的研究等 [18] 。此外,在人工智慧領域,多智慧體、人工集體智慧的研究,也可以歸結為人工生態系和社會的研究 [19] 。
開放式演化
「除了演化的光輝,生物學中的一切都沒有意義。」
——美國前進演化生物學家狄奧多西·多布贊斯基(Theodosius Dobzhansky)
在差異性選擇的驅動下,進行繁殖、遺傳和變異的生物族群和生態系,會經歷達爾文式的演化。前進演化演算法 [20] 使用類似的概念來搜尋外部環境適應度函式(fitness)的最優值,往往會帶來令人意想不到的結果 [21] 。
然而,自然演化在多個層面上要比人工模擬復雜得多,包括基因組復雜性、族群規模、世代數量、從水平基因轉移到性選擇的繁殖策略、基因調控、發育和表觀遺傳學的作用、多個共同演化族群之間的互動以及內在定義的適應度等方面。這些更自然的演化特征構成了ALife演化研究的核心內容 [14][22] 。
如達爾文言,「無盡之形最美」 [23] , 開放式演化 (Open-Ended Evolution, OEE) [24] ,是ALife的一個重要研究領域。它不僅捕捉了現有生命系統各種內容、包括與環境作用關系的根本特征,同時提供了一個框架來探索和模擬潛在生命形式的可能性空間。在研究過程中,這有助於啟發各種最佳化、學習和前進演化等演算法。因此, 實作開放式演化也可以說是人工生命的終極目標。
區別於一般演化,一個開放式演化系統永遠不會陷入單一的穩定平衡,其新穎性會持續產生 [25] ,復雜性會無限增加。這表明開放式演化是一個表征了多個尺度演化的過程 [26] 。開放式演化領域涉及生命起源問題 [27] 、復雜性和組織層次出現的重大演化轉變 [28] 以及元演化,即演化能力及其演化的演化 [29] 等問題。
要產生開放式演化,有幾個假設條件 [30] :
(1)潛在基因型的無限遺傳空間。 這並不意味生物體的基因組長度必須沒有限制地變化。由於主導基因的突變和遺傳速度很慢,也可能存在大量非編碼蛋白的調控基因,即原來所謂的垃圾基因,這些基因透過調控前者可以迅速產生多樣性。這意味著只需要透過改變類似於神經網路中權重的調控開關,可能很快就能形成新的物種。這一假設可以幫助解釋普遍共同祖先(LUCA)的生物共同基因,以及寒武紀生物大爆發。
(2)潛在表型之間存在多種突變途徑。 這意味著潛在生物體特征應該能夠由許多不同的突變途徑產生,例如人和章魚的眼睛雖功能相似,但卻是獨立前進演化的;魚和海豚的鰭足則是趨同演化的案例。理論生物學家史超活·考夫曼(Stuart Kauffman)透過隨機布爾網路對此進行了刻畫,他發現基因調控網路的最終行為是由節點數量以及各節點的入度K決定的。當K=2時,網路就會進入既不是不動點、振蕩 ,也不是完全混沌的混沌邊緣的狀態。這時網路每一種吸子就對應一種細胞的基因表現,即不同的基因組,可以采取了不同的前進演化路徑來達到與環境適配的同一目標表型。
(3)持續演化的動態自適應景觀 (Dynamic adaptive landscape)。這意味著生物體周圍的環境是動態變化的。一方面,隨著生物族群演變,其周圍的環境也隨其行為而改變,一方面動態的環境會不斷對生物族群進行選擇、透過表觀遺傳等作用影響基因型和表型的實作。
開放式演化可以在元胞自動機,數位前進演化模擬平台如Tierra、Polyworld、Avida等,人工化學 [31] 模擬平台Stringmol等 [32] 進行研究。2019年一篇研究 [33] ,根據演化的表型行為與搜尋空間的關系,確定了三種型別的新穎性以及對應的開放性:
探索開放性(Exploratory): 可使用當前模型描述的新穎性,通常涉及現有元件的重新組合或現有參數值的修改。例如基因組中新的等位基因的組合可能決定了新脊椎動物的脊椎數目。
擴充套件開放性(Expansive): 一種需要改變模型,但仍使用當前元模型中存在的概念的新穎性,涉及在狀態空間中發現開啟新鄰域的狀態。這往往涉及組織利用先前未被利用的化學或物理規律,或一個新的邊界條件或裝置的出現。例如飛行翅膀的出現,以及視覺感官系統的出現。
變革開放性(Transformational): 一種引入新概念、需要改變元模型的新穎性。這不僅涉及利用新的物理定律,還包括了組織層級上的變遷。例如,合成一種以前未在代謝反應中使用的新化學物質。生命起源、真核細胞以及多細胞生物、大腦和自我意識、文字和技術都屬於此類。這往往對應於復雜系統中的強湧現。
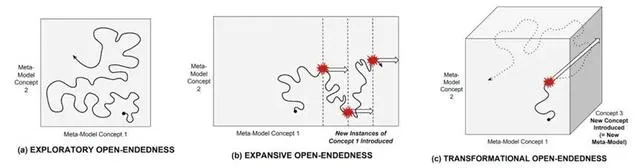

▷ 三種開放性:達爾文雀對生態席位的適應;翅膀出現;真核細胞。
同樣,對開放式演化有幾種不同的度量指標也用於檢測 [34] 或量化其開放性潛力,例如前進演化動態測量(MODES)指標 [35] 。在NK模型和Avida數位前進演化平台等不同的實驗環境中,這些指標的有效性和適用性已經得到驗證。
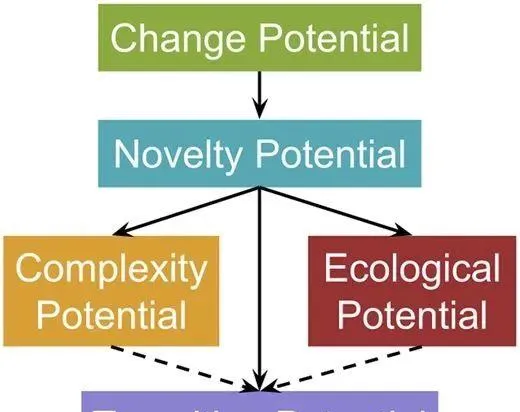
▷ MODES各指標之間的關系。
在 MODES 中,復雜性潛力需要機體能夠不斷地將更多的環境資訊整合到自身基因組中,從而產生更復雜的行為,生態性潛力描述的是族群作為一個整體能夠吸收和反映環境資訊的能力,包括生物與非生物環境交互作用創造新的生態席位和營養層次的能力。
可見,要實作持續的開放性變革,必須將環境和生態因素納入生物的演化過程中。據此,開放式演化系統可以視為Rosen的(M,R)系統。此外,據大語言模型ChatGPT描述,它更傾向於探索開放性。開放式演化的研究勢必會對計算創造力和AGI研究提供重要的方法和範式。
共生
「演化中的一切,只有在寄生的光芒下才有意義」,這篇2021年一篇論文的標題 [36] ,模仿了多布贊斯基對演化的描述,但這或許並不誇張。諸多研究表明,寄生-互利共生連續譜的演化 [37] ,是生成生物新穎性、塑造生態多樣性和推動地球生命重大轉變的重要機制。如前所述,尼爾斯·貝瑞切利的人工生命模擬早期就洞悉了這點。尼克·萊恩(Nick Lane)在【復雜生命的起源】中也認為,內共生提高了生命能量利用效率,促進了真核生物的起源,並使其最終演化成地球復雜生命。
共生(Symbiosis)可以看做是生物體和族群之間,在更多樣的生態尺度下更為密切的協同演化 [38] 的結果。根據空間位置,共生可以分為內共生(ectosymbiosis)和外共生(endosymbiosis),根據物種間的利益關系,則通常包括互利共生、片利共生、中性互作、偏害共生、競爭以及寄生 [39] /捕食(廣義) [40] 。生態系中資源交換-利用依賴迴圈會導致社群構建和共生演化,甚至形成跨餵養(crossfeeding)現象,即一個物種的代謝物成為另一個物種的資源 [41] 。
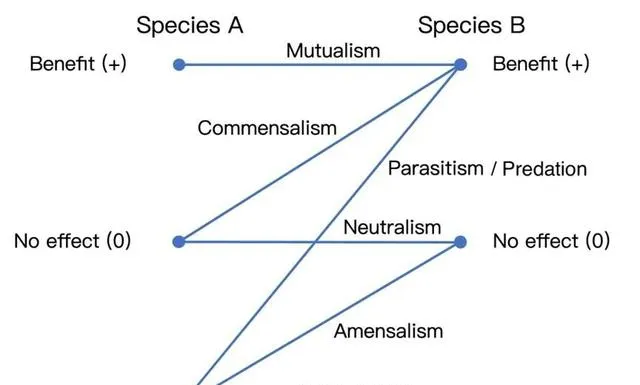
▷ 伴侶關係 [42] 。作者結合 https://en.wikipedia.org/wiki/Symbiosis以及[40] Figure 1.1圖表合並自制。
共生同樣可以用生命遊戲、數位前進演化模擬平台進行研究。例如,有研究者在康威的生命遊戲基礎上發展了Model-S模型 [43] ,該模型成功模擬產生了自組織、自創生、多細胞的演化、有性生殖的誕生以及物種共生的融合策略。研究發現,即使是少量共生也能顯著提高族群的適應度,並可能支持開放式演化。在支持共生的數位前進演化系統中 [44] ,數位生物透過消耗「CPU時間」資源來生存和發展,內共生物則消耗宿主內部資源,與真實世界中細胞的粒線體類似。同樣,數位核糖體將數位基因組(二進制程式碼)轉化(代謝)為某種輸出或行為,類似於生物細胞中核糖體讀取遺傳程式碼並合成蛋白質的過程。
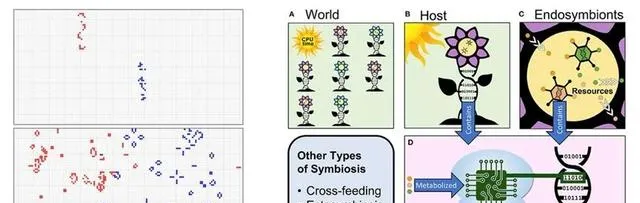
▷ Model-S模擬的移民遊戲產生「共生層」 [43] 與數位前進演化系統中模擬的內共生 [44] 。
可以說, 生命從起源起就是一種共生現象,並且橫跨了不同種類的復制子。從基因到文化(模因),到以技術作為擴充套件表型(技因),未來,生物與數位的融合可能進入一個人機共生的新時代 [45] 。
如果生物演化的基質基於自然物理環境,那麽技術物-數位物的演化基質和生態環境又是什麽?顯然,這必須是一個人類和機器生命都能進行互操作的系統。有研究指出 [46] ,基於區塊鏈的分布式系統具有對環境響應、生長和變化、遺傳復制和達到穩態等諸多符合生命定義的特征。若和神經網路等AI技術結合,這種系統將成為比傳統生命更有優勢的自組織系統。文章認為,基於公鏈的分布式虛擬機器(dVM)提供了一個自主前進演化能力的開放環境,為通用人工智慧(AGI)的發展提供支持。
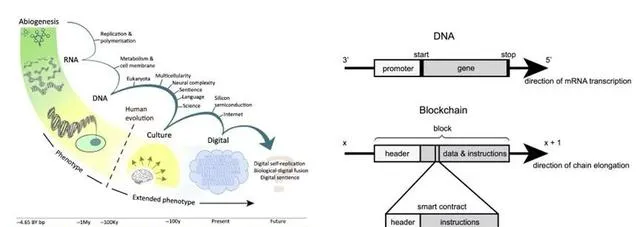
▷ 前進演化基質的演化 [45] ;DNA與區塊鏈 [46] 。
最近,區塊鏈技術領域興起了一種名為數位物質理論(Digital Matter Theory)的觀點, 該理論認為,數位資訊可以被看作一種數位物質,與實體物質(比如木頭或金屬)類似。 透過挖掘區塊數據中固有的模式,可以建立一種新型的數位物質形態,即區塊元素表。它被視為數位世界物質基質和資產的一種表現。隨著區塊高度增加,這個數位世界也和物質世界一樣在不斷生長,其發展同樣不可預期。在這樣的系統中,人們可以發行非任意同資源/代幣(Non-Arbitrary,意味著總量非人為設定或隨意增發),為自主世界與元宇宙提供了底層邏輯和基礎。
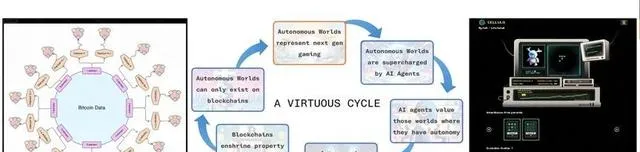
▷ 數位物質理論、自主世界和鏈上生命遊戲Cellula [55] 。
此外,目前也出現一些基於區塊鏈的Alife計畫。例如鏈上的人工生命模擬遊戲(Cellula),它允許玩家自由地建立各種基因序列,並孕育出自己的鏈上「生命」,在這個遊戲中,區塊高度扮演了「時間」的角色,每個「生命」都會在鏈上時空生態系中成長、前進演化、死亡。區塊鏈上的Alife,由於涉及到了玩家和鏈上智慧體主體,存在註意力流和資訊的雙向傳遞,因此是一種典型的互惠共生形態。此外,還有一些計畫探索了透過區塊鏈技術或物理環境,使非人類主體如植物與數位或機器共生的可能性,例如terra0和 Flora Robotica計畫。

▷ 跨媒介跨物種共生的例子。
當然,比較廣泛討論的人機共生是人和人工智慧方面和共生。這主要基於任務和協作的研究、以及人機價值對齊 [47] 、尤其AI心智理論 [48] 方面的研究。這些研究在很大程度上體現了人類中心主義的觀點以及以人類利益為主的共生設計考量。
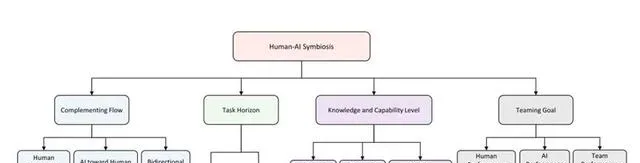
▷ 人機共生的方法維度。 [49]
生物啟發AI與人工生命的未來
回顧歷史可以看到,AI曾長期受制於符號人工智慧桎梏,即計算啟發方法(GOFAIstic) [50] ,而 Alife和生物啟發方法 (亦被稱為控制論啟發) 則為AI的發展提供了重要靈感和範式。
辛頓(Geoffrey Hinton)開創的深度學習網路,實際上是一種模擬人腦神經的AI系統,而像Transformer這樣的大模型透過自註意力機制在某種程度上模擬了人類大腦的記憶和學習機制,即人腦的海馬體記憶模型是一個帶了迴圈神經網路的因果Transformer(「TEM is causal transformer with RNN position encodings」) [51] 。開發出 Chatgpt 的OpenAI,時刻提醒員工「苦澀的教訓」(The bitter lesson),堅信規模法則(Scaling Law)就能湧現內容(Emerging properties) ,認為只要持續不斷增大數據和算力的大模型才能實作AGI。強化學習包括人類反饋強化學習(RLFH),源自對動物行為心理學的借鑒,本質是一種基於反饋的環境適應機制,生成式對抗網路(GAN)則源自動物競爭與共生的生態學機制。
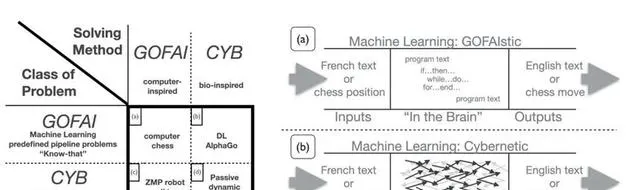
▷ GOFAI主義與生物啟發(控制論)AI [52]
隨著技術進步,Alife研究者不斷探索從抽象模型到計算模擬,再到具身嵌入、人工生態、開放式演化等多種範式,這些都在推動AI的進一步發展。例如,卡爾·弗裏斯頓(Karl Friston)的自由能原理和楊立昆的聯合嵌入預測架構(I-JEPA) [53] 都嘗試在AI系統中整合更多生物學原理,以增強其與環境的互動能力和自主性。甚至包括非馮諾依曼架構、辛頓後來提出的可朽計算(Mortal Computation)、如今廣為討論的人機價值對齊,以及受生態啟發的人工智慧 [56] 等研究,都可以看到Alife的發展進路的影子。
當然, 目前的生成式AI,特別是大語言模型,也在對Alife研究產生深遠影響 [54] 。例如利用大語言模型探索意義的產生、湧現和因果湧現、開放式演化,尤其是人機協作、人工集體智慧以及大語言模型驅動的多AI智慧體系統(著名的史丹佛AI小鎮)。此外,目前也有演算法層面的結合,例如結合神經網路和遺傳演算法的神經前進演化(Neuroevolution),已有研究者使用簡單的虛擬生物 [57] ,透過視覺、聽覺感知環境,並與同類和捕食者互動,發現部份個體湧現出專門應對捕食者的"恐懼中樞"模組,這表明類似生物腦的情緒和意識等神經機制可以在人工基質中湧現。
最後,如果我們以Alife的範式去思考,我們面前展開的問題有:基於區塊鏈的系統和自主世界等數位原生基質如何實作具有自主性的(M,R)系統?大語言模型和AI智慧體如何演化出變革開放性?AI和人類將會是形成哪種伴侶關係,競爭、內共生還是跨物種餵養?虛擬現實和擴充套件現實又將如何影響未來的數位生態?
不管未來如何,Alife、AI、區塊鏈和元宇宙(AR/VR/XR)將相互交織發展,帶來一個自然生命和虛擬主體、數位和物理的相融的共生世界。在這樣的後人類時代,不管是哪種形式,生命實體都將在廣闊無垠的宇宙中繼續其開放演化的旅程。
參考文獻:
[1] D. B. Fogel, "Nils Barricelli - artificial life, coevolution, self-adaptation," in IEEE Computational Intelligence Magazine, vol. 1, no. 1, pp. 41-45, Feb. 2006, doi: 10.1109/MCI.2006.1597062.
[2] Alan Dorin, Susan Stepney; What Is Artificial Life Today, and Where Should It Go?. Artif Life 2024; 30 (1): 1–15. doi: https://doi.org/10.1162/artl_e_00435
[3] https://medium.com/paul-austin-murphys-essays-on-philosophy/christopher-langton-artificial-life-and-ultra-functionalism-3eb5fa8a92f0
[4] (M,R)即代謝(Metabolism)和修復(Repair)。代謝由一組機制(記為f)完成從材料A到產物B的轉化,修復則由一組機制(記為Φ)從代謝產物B合成代謝機制f本身。
[5] Maturana, H. R., & Varela, F. J. (1980). Autopoiesis and Cognition: The Realization of the Living. D. Reidel Publishing Company.https://doi.org/10.1007/978-94-009-8947-4
[6] Dolson E and Ofria C (2021) Digital Evolution for Ecology Research: A Review. Front. Ecol. Evol. 9:750779. doi: 10.3389/fevo.2021.750779
[7] Pattee, H.H. (2012). Evolving Self-reference: Matter, Symbols, and Semantic Closure. In: LAWS, LANGUAGE and LIFE. Biosemiotics, vol 7. Springer, Dordrecht. https://doi.org/10.1007/978-94-007-5161-3_14
[8] Stepney, S. (2007). Embodiment. In D. Flower & J. Timmis, (Eds.) In silico immunology (pp. 265–288). Springer. https://doi.org/10.1007/978-0-387-39241-7_13
[9] https://towardsdatascience.com/robot-farms-artificial-life-and-second-order-engineering-an-in-depth-interview-with-professor-a524c92ee455
[10] Reiners, William A. 「Complementary Models for Ecosystems.」 The American Naturalist, vol. 127, no. 1, 1986, pp. 59–73. JSTOR, http://www.jstor.org/stable/2461647. Accessed 27 May 2024.
[11] Lindgren, K., & Nordahl, M. G. (1994). Artificial food webs. In C. G. Langton (Ed.), Artificial Life III: Proceedings of the workshop on Artificial Life, Santa Fe, 1992 (pp. 73–103). Addison-Wesley.
[12] Theraulaz, G., & Bonabeau, E. (1999). A brief history of stigmergy. Artificial Life, 5(2), 97–116. https://doi.org/10.1162/106454699568700
[13] Luo, T.-T., Heier, L., Khan, Z. A., Hasan, F., Reitan, T., Yasseen, A. S., III, Xie, Z.-X., Zhu, J.-L., & Yedid, G. (2018). Examining community stability in the face of mass extinction in communities of digital organisms. Artificial Life, 24(4), 250–276.
[14] Bull, L. (2021). On the emergence of intersexual selection: Arbitrary trait preference improves female-male coevolution. Artificial Life, 27(1), 15–25. https://doi.org/10.1162/artl_a_00335
[15] Dorin, A., Korb, K. B., & Grimm, V. (2008). Artificial-life ecosystems: What are they and what could they become? In S. Bullock, J. Noble, R. Watson, & M. A. Bedau, (Eds.) Artificial Life XI (pp. 173–180). MIT Press.
[16] H. Hamann et al., "Flora Robotica - Mixed Societies of Symbiotic Robot-Plant Bio-Hybrids," 2015 IEEE Symposium Series on Computational Intelligence, Cape Town, South Africa, 2015, pp. 1102-1109, doi: 10.1109/SSCI.2015.158.
[17] Marriott, C., Borg, J. M., Andras, P., & Smaldino, P. E. (2018). Social learning and cultural evolution in Artificial Life. Artificial Life, 24(1), 5–9. https://doi.org/10.1162/ARTL_a_00250
[18] Yamauchi, H., & Hashimoto, T. (2010). Relaxation of selection, niche construction, and the Baldwin effect in language evolution. Artificial Life, 16(4), 271–287. https://doi.org/10.1162/artl_a_00006,
[19] https://arxiv.org/abs/2304.05147
[20] De Jong, K. (2005). Genetic algorithms: A 30-year perspective. In L. Booker, S. Forrest, M. Mitchell, & R. Riolo, (Eds.) Perspectives on adaptation in natural and artificial systems (pp. 1–32). Oxford University Press. https://doi.org/10.1093/oso/9780195162929.003.0006
[21] Lehman, J., Clune, J., Misevic, D., Adami, C., Beaulieu, J., Bentley, P. J., Bernard, S., Belson, G., Bryson, D. M., Cheney, N., Cully, A., Donciuex, S., Dyer, F. C., Ellefsen, K. O., Feldt, R., Fischer, S., Forrest, S., Frénoy, A., Gagneé, C., … Yosinksi, J. (2020). The surprising creativity of digital evolution: A collection of anecdotes from the evolutionary computation and Artificial Life research communities. Artificial Life, 26(2), 274–306. https://doi.org/10.1162/artl_a_00319
[22] Liard, V., Parsons, D. P., Rouzaud-Cornabas, J., & Beslon, G. (2020). The complexity ratchet: Stronger than selection, stronger than evolvability, weaker than robustness. Artificial Life, 26(1), 38–57. https://doi.org/10.1162/artl_a_00312
[23] Darwin, C. R. (1872). On the origin of species (6th ed.). Dent. https://doi.org/10.5962/bhl.title.28875
[24] https://alife.org/encyclopedia/introduction/open-ended-evolution/
[25] Soros, L., & Stanley, K. (2014). Identifying necessary conditions for open-ended evolution through the artificial life world of Chromaria. In H. Sayama, J. Rieffel, S. Risi, R. Doursat, & H. Lipson (Eds.), ALIFE 14: Proceedings of the Fourteenth International Conference on the Synthesis and Simulation of Living Systems (pp. 793–800). MIT Press. https://doi.org/10.7551/978-0-262-32621-6-cp28
[26] Corominas-Murtra, B., Seoane, L. F., & Solé, R. (2018). Zipf’s Law, unbounded complexity and open-ended evolution. Journal of The Royal Society Interface, 15(149), 20180395. https://doi.org/10.1098/rsif.2018.0395
[27] Cardoso, S. S. S., Cartwright, J. H. E., Čejková, J., Cronin, L., De Wit, A., Giannerini, S., Horváth, D., Rodrigues, A., Russell, M. J., Sainz-Díaz, C. I., & Tóth, Á. (2020). Chemobrionics: From self-assembled material architectures to the origin of life. Artificial Life, 26(3), 315–326. https://doi.org/10.1162/artl_a_00323
[28] Maynard Smith, J., & Szathmáry, E. (1995). The major transitions in evolution. Oxford University Press
[29] Payne, J. L., & Wagner, A. (2019). The causes of evolvability and their evolution. Nature Reviews Genetics, 20(1), 24–38. https://doi.org/10.1038/s41576-018-0069-z
[30] Taylor, T. (2012). Exploring the Concept of Open-Ended Evolution. https://www.tim-taylor.com/papers/taylor2012exploring.pdf
University of New Mexico, Albuquerque, NM 87131, Ackley, D., & Small, T. (2014). Indefinitely Scalable Computing = Artificial Life Engineering. Artificial Life 14: Proceedings of the Fourteenth International Conference on the Synthesis and Simulation of Living Systems, 606–613. https://doi.org/10.7551/978-0-262-32621-6-ch098
[31] Banzhaf, W., & Yamamoto, L. (2015). Artificial chemistries. MIT Press. https://doi.org/10.7551/mitpress/9780262029438.001.0001
[32] Hickinbotham, S., Clark, E., Stepney, S., Clarke, T., Nellis, A., Pay, M., & Young, P. (2011). Molecular microprograms. In ECAL 2009, Budapest, Hungary (pp. 297–304). Springer. https://doi.org/10.1007/978-3-642-21283-3_37
[33] Tim Taylor; Evolutionary Innovations and Where to Find Them: Routes to Open-Ended Evolution in Natural and Artificial Systems. Artif Life 2019; 25 (2): 207–224. doi: https://doi.org/10.1162/artl_a_00290
[34] Susan Stepney, Simon Hickinbotham; On the Open-Endedness of Detecting Open-Endedness. Artif Life 2023; doi: https://doi.org/10.1162/artl_a_00399
[35] Emily L. Dolson, Anya E. Vostinar, Michael J. Wiser, Charles Ofria; The MODES Toolbox: Measurements of Open-Ended Dynamics in Evolving Systems. Artif Life 2019; 25 (1): 50–73. doi: https://doi.org/10.1162/artl_a_00280
[36] Hickinbotham Simon J., Stepney Susan and Hogeweg Paulien 2021Nothing in evolution makes sense except in the light of parasitism: evolution of complex replication strategiesR. Soc. Open Sci.8210441
[37] Vostinar, A. E., & Ofria, C. (2018). Spatial structure can decrease symbiotic cooperation. Artificial Life, 24(4), 229–249. https://doi.org/10.1162/artl_a_00273
[38] Weisbuch, G., Duchateau, G. Emergence of mutualism: Application of a differential model to endosymbiosis. Bltn Mathcal Biology 55, 1063–1090 (1993). https://doi.org/10.1007/BF02460700
[39] Hickinbotham, S., Stepney, S., & Hogeweg, P. (2021). Nothing in evolution makes sense except in the light of parasitism: Evolution of complex replication strategies. Royal Society Open Science, 8(8), 210441. https://doi.org/10.1098/rsos.210441
[40] Bronstein, Judith L. (ed.), 'The study of mutualism', in Judith L. Bronstein (ed.), Mutualism (Oxford, 2015; online edn, Oxford Academic, 17 Sept. 2015), https://doi.org/10.1093/acprof:oso/9780199675654.003.0001
[41] Crombach A, Hogeweg P. Evolution of resource cycling in ecosystems and individuals. BMC Evol Biol. 2009 Jun 1;9:122. doi: 10.1186/1471-2148-9-122. PMID: 19486519; PMCID: PMC2698886.
[42] 作者結合 https://en.wikipedia.org/wiki/Symbiosis 以及[40] Figure 1.1圖表合並自制.
[43] Turney PD. Evolution of Autopoiesis and Multicellularity in the Game of Life. Artif Life. 2021 Jun 11;27(1):26-43. doi: 10.1162/artl_a_00334. PMID: 34529755.
[44] Vostinar AE, Skocelas KG, Lalejini A and Zaman L (2021) Symbiosis in Digital Evolution: Past, Present, and Future. Front. Ecol. Evol. 9:739047. doi: 10.3389/fevo.2021.739047
[45] Gillings MR, Hilbert M, Kemp DJ. Information in the Biosphere: Biological and Digital Worlds. Trends Ecol Evol. 2016 Mar;31(3):180-189. doi: 10.1016/j.tree.2015.12.013. Epub 2016 Jan 8. PMID: 26777788.
[46] Abramov, O., Bebell, K.L. & Mojzsis, S.J. Emergent Bioanalogous Properties of Blockchain-based Distributed Systems. Orig Life Evol Biosph 51, 131–165 (2021). https://doi.org/10.1007/s11084-021-09608-1
[47] https://arxiv.org/abs/2310.19852
[48] https://arxiv.org/abs/2202.09447
[49] https://arxiv.org/pdf/2103.09990
[50] https://zh.wikipedia.org/wiki/GOFAI
[51] https://arxiv.org/abs/2112.04035
[52] Harvey I. Motivations for Artificial Intelligence, for Deep Learning, for ALife: Mortality and Existential Risk. Artif Life. 2024 Feb 1;30(1):48-64. doi: 10.1162/artl_a_00427. PMID: 38346273.
[53] https://arxiv.org/abs/2301.08243
[54] https://direct.mit.edu/artl/article/29/2/141/116359/Editorial-What-Have-Large-Language-Models-and
[54]https://doi.org/10.1162/isal_a_00431
[55] https://medium.com/p/68c29c1e374e
https://medium.com/p/2f1853f8a1d7
https://medium.com/p/1d81efda13;
[56] Han BA, Varshney KR, LaDeau S, Subramaniam A, Weathers KC, Zwart J. A synergistic future for AI and ecology. Proc Natl Acad Sci U S A. 2023 Sep 19;120(38):e2220283120. doi: 10.1073/pnas.2220283120. Epub 2023 Sep 11. PMID: 37695904; PMCID: PMC10515155.
[57] Kevin Godin-Dubois, Sylvain Cussat-Blanc, Yves Duthen; July 18–22, 2021. "Spontaneous modular NeuroEvolution arising from a life/dinner paradox." Proceedings of the ALIFE 2021: The 2021 Conference on Artificial Life. ALIFE 2021: The 2021 Conference on Artificial Life. Online. (pp. 95). ASME. https://doi.org/10.1162/isal_a_00431











